
Addisu Sileshi
Jimma University College of Agriculture and Veterinary Medicine, P.O. Box 307, Jimma, Ethiopia
Waktole Sori
Jimma University College of Agriculture and Veterinary Medicine, P.O. Box 307, Jimma, Ethiopia
Mohamed Dawd
Ethiopian Institute of Agricultural Research, Ambo Plant Protection Research Center, P.O.Box 2003, Addis Ababa, Ethiopia
Asian Journal of Plant Sciences
Year: 2013 | Volume: 12 | Issue: 1 | Page No.: 1-10







ABSTRACT
Termites, Macrotermes are major agricultural and domestic problem in Ethiopia causing serious damage with loss up to 100%. The use of synthetic termitecids was the most commonly used prevention measure to reduce the termites attack. However, these synthetic termiticides were known to be very harmful to the environment and non-target organisms. Therefore, efficacy of entompathogenic fungi, Metarhizium anisopliae (isolates PPRC-2 and MM) and Beauveria bassiana (isolates PPRC-56 and 9609) were evaluated against Macrotermes. The isolates were obtained from Ambo Plant Protection Research Center, Ethiopia. For each isolate concentrations of 1x105 to 1x109 conidia mL-1 were prepared and used as treatments. Untreated and standard (Diazinon 60% EC) checks were used for comparison. The treatments were laid out in Completely Randomized Design (CRD) and replicated thrice. The fungal isolates were evaluated by direct spraying of spore suspensions on worker Macrotermes spp. The result of the study revealed that all fungal isolates used were able to infect and cause mortality at all concentrations. The percent mortality of Macrotermes varied from 60 to 100% for M. anisopliae isolate MM at 1x105 to M. anisopliae isolate PPRC-2 at 1x109, respectively. Similarly, the percentage mortality of Macrotermes varied from 25-95% for B. bassiana isolate 9609 at low concentration and isolate PPRC-56 at highest concentration, respectively. The isolates had LT50 ranging from 7.74 days in M. anisopliae isolate PPRC-2 to 8.80 days in B. bassiana isolate 9609. The concentration response with the isolate PPRC-2 showed the lowest LC50 of 3.21x105 conidia mL-1 followed by isolates MM, PPRC-56 and 9609 with LC50 of 3.82x105, 4.39x105 and 5.08x105 conidia mL-1, respectively. In conclusions, the present study suggests that the use of entompopathogenic fungi, M. anisopilae and B. bassiana, at higher concentrations for seven days is an eco-friendly effective mycoinsecticides that causes more than 95% mortality of Macrotermes.
PDF Abstract XML References Citation
Received: December 05, 2012;
Accepted: March 11, 2013;
Published: June 21, 2013
How to cite this article
Addisu Sileshi, Waktole Sori and Mohamed Dawd, 2013. Laboratory Evaluation of Entomopathogenic Fungi Metarhizium anisophilae
and Beauveria bassiana Against Termite, Macrotermes (Isoptera: Termitidae). Asian Journal of Plant Sciences, 12: 1-10.
DOI: 10.3923/ajps.2013.1.10
URL: https://scialert.net/abstract/?doi=ajps.2013.1.10
DOI: 10.3923/ajps.2013.1.10
URL: https://scialert.net/abstract/?doi=ajps.2013.1.10
INTRODUCTION
Termites belong to the insect order Isoptera and have long been recognized as important agricultural and domestic pests (Owusu et al., 2008; Samb et al., 2011). Among all termites, the largest termite species are the Macrotermes spp. known to be fungus growing and mound building. Macrotermes builds large epigial nests (mounds) from where they forage outwards to distances up to 50 m in galleries (Osipitan and Oseyemi, 2012). Macrotermes are economically important insect pests causing serious damage up to 100% loss to agricultural crops and various domestic products (Abdurahman, 1990; UNFAO, 2000; Sekamatte, 2001; Nyeko et al., 2010). Damage by these termite species to agricultural and commercial products also provides entry for secondary infection by pathogen especially Asperagilous, which cause indirect yield loss and contamination of products with aflatoxins (Osipitan and Oseyemi, 2012).
Management of termites has largely relied on broad spectrum and persistent organo chlorine insecticides in the world (Logan et al., 1990). Nowadays, most of the persistent insecticides were banned or withdrawn from the market for human health and environmental reasons. Also, synthetic insecticides are not affordable by poor farmers of Africa including Ethiopia. Thus, there were serious limitations and increasing legal restrictions associated with the application of persistent and deleterious insecticides; because of which the search for environmentally benign alternative methods of termite management has been intensified by entomologists. Among the diverse potential alternatives available for termite management, the use of entomopathogenic fungi is getting momentum (Michael, 2005).
Entomopathogenic fungi, Beauveria bassiana (Basamo) and Metarhizium anisophilae (Metchnikoff) are effective in the management of different species of termite (Milner et al., 1998; Milner, 2003) which may be used in different methods among which direct exposure, soil barrier and in baits system were able to achieve good control in termite colony. The genus Metarhizium, Beauveria and Paecilomyces are fungal pathogens of insects that have shown great promise in commercial development. M. anisopliae, with worldwide distribution has been isolated from more than 200 insect species across seven orders and has shown great potential as bio-control agents. Strains of Metarhizium differ in their host range, necessitating selection of the most virulent against a target insect (Zimmermann, 1993).
M. anisopliae has an advantage over B. bassiana in microbial management of termite due to their social behavior and high production of fungal biomass (Sun et al., 2002). Pik-Kheng et al. (2009) reported that isolates of M. anisopliae were pathogenic against subterranean termite, Coptotermes curvignathus, causing 100% mortality at 1x107 conidia mL-1 within 3 days post-inoculation. According to David et al. (2010), a positive relationship between virulence and repellency effect of different isolates of the fungus M. anisopliae on M. michaelseni was determined. Further, they compared the volatile profiles of two isolates of M. anisopliae, on the same species of termite and found that the volatiles of each isolate act synergistically.
The entomopathogenic fungus, M. anisophilae, has more host ranges and is widely used as bio-pesticide agents on several types of insect pests which include onion thrips, storage cowpea, white grub, cattle ticks and different species of termites such as Reticulitermes spp., Rhinotermitidae, Coptotermes formosanus and Odontotermes formosanus (Maniania et al., 2003; Sun et al., 2003; Wang and Powell, 2004; Cherry et al., 2005; Samson et al., 2005; Bahiense et al., 2006; Dong et al., 2009). Many strains of M. ansisopliae have been isolated from termites and are reported as effective myco-insecticides (Sun et al., 2003; Wright et al., 2005) for the management of subterranean termites. Entomopathogenic fungi isolate, Aspergillus terreus, spore suspension at different concentration was tested on different stage of tick, Hyalomma anatolicum anatolicum and has shown promising result as bio-pesticides (Suliman and Mohammed, 2012).
Biological control of different agricultural and domestic pests has been reported as promising option in the current and future state of pest management (Bittencourt, 2000; Chandler et al., 2000; Kaaya, 2003; Sharma et al., 2011; Jaramillo and Borgemeister, 2006). Entomopathogenic fungi, M. anisophilae, pathogenicity has been studied and found to cause mortality on different stage of insect pests such as filth fly parasitoid, Spalangia Cameroni, Malaria mosquito, Anopheles Gambia (Nielsen et al., 2004; Scholte et al., 2006). Thus new management approach in Macrotermes is deemed of prime necessity in Ethiopia. However, the report on the use of entomopathogenic fungi for the management of termite in Ethiopia is very meager despite the economic importance of Macrotermes as insect pest. Therefore, the current study was aimed at determining the efficacy of some available isolates of entomopathogenic fungi (B. bassiana and M. anisophilae) at different concentrations for the management of Macroterms.
MATERIALS AND METHODS
Description of the experimental area: The experiment was conducted in Entomology and Plant Pathology Laboratory, Jimma University, College of Agriculture and Veterinary Medicine (JUCAVM) which is located at 354 km Southwest of Addis Ababa, Ethiopia. JUCAVM is at geographical coordinate of 7°42’ N latitude and 36°50’ E longitude with an altitude of 1710 masl. The temperature ranges from 11.8°C (minimum) to 26.8°C (Maximum) with maximum relative humidity and mean rainfall of 91% and 1500 mm per annum, respectively (Abera et al., 2011).
Termite collections and establishment for test: Termite, Macrotermes spp., population were collected from two main sites, namely Saye Kebele of Mana and near Agaro town of Agro Woredas. At these two localities, termite mounds were dug up using spade and soil containing termites were put on plastic sheets. Termites were collected from the plastic sheets using camel hair brush and placed in plastic boxes (polyethylene plastic box) (2217x7 cm3) as described by Gitonga et al. (1995). Wooden plants (termite infested materials) were added to the plastic boxes as feed for the termites. Then the top parts of the plastic boxes were covered with a mesh cloth that allows air ventilation in and out easily but preventing the escape of the termites. Moistened wad of cottons were placed in the plastic boxes to maintain the required moisture level (more than 60%) for the survival of termites. The boxes carrying the termites were transported to JUCAVM, Entomology and Plant Pathology Laboratory and placed in a cool and dark area until needed for the experiments. Periodically, dry wooden materials were provided to the termite’s population and the plastic boxes were inspected for maintenance of the required moisture level.
Source of entomopathogenic fungi isolates: Two already cultured species of entomopathogenic fungi (M. anisopilae and B. bassiana) and two isolates of each species, viz., MM, PPRC-2 (M. anisopliae) and 96O9 and PPRC-56 (B. bassiana) used in this experiment were obtained from Ethiopian Institutes of Agricultural Research, Ambo Plant Protection Research Center.
Preparation of spore suspension: To prepare fungal spore suspension, the conidia were harvested by flooding each plate with 10 mL of 0.05% Tween 80 (sigma) as a wetting agent in sterile distilled water. The conidial suspensions were prepared by mixing the solution using a magnetic stirrer for 5 min and the suspensions were filtered through sterile muslin cloth to eliminate the coagulated medium. The spore suspension of 1x105, 1x106, 1x107, 1x108 and 1x109 per mL of distilled water were prepared for each isolates by using haemocytometer as described by Lacey (1997).
Experimental design and treatments: There were 22 treatments (Table 1) replicated thrice and laid-out in Completely Randomized Design (CRD). In this experiment, there were two controls (the untreated and the standard controls, viz., sterile water and 0.21% a.i. Diazinon, respectively) for comparison. Each isolate was tested at five concentrations (1x105, 1x106, 1x107, 1x108 and 1x109 conidia mL). The experiments were conducted under laboratory conditions at 25±2°C temperature and 60% RH in dark places.
Median Lethal concentration (LC50) and time (LT50): Median lethal time (LT50) and Median lethal concentration (LC50) were determined for each concentration by taking in to account the time and concentration required at which the inoculums of fungus caused 50% of the mortality on worker termite population.
| Table 1: | Entomopathogenic fungi isolates investigated in the present study |
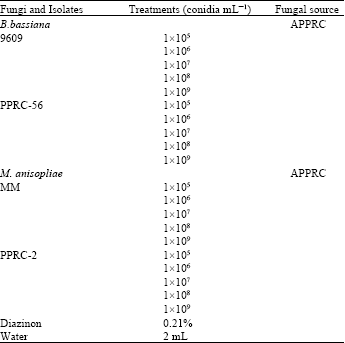 | |
| APPRC: Ambo Plant Protection Research Center | |
Lethal time (LT50) and lethal concentration (LC50) required to achieve 50% mortality per replicate was obtained from probit analysis.
Mortality of termite due to fungal isolates: To test the efficacy of each of the fungal isolates on termites, twenty worker termites were placed in a Petri dish on a filter paper and sprayed with the spore solutions as described by Julia and Lina (2010). To determined how many termites were dead without being infested with entomopathogenic fungi, a control group were sprayed with only sterile distilled water as a negative control. The efficacy was evaluated on a daily basis for 15 days, by counting dead termites which were later converted to percentage mortality. Petri dishes containing the treated termites were maintained at 25±2°C in the dark. Mortality data were corrected for the corresponding control mortality by the formula:
where, CM is corrected mortality, T is mortality in treated insects and C is mortality in untreated insects (Abbott, 1925).
Data analysis: The data recorded for different response variables in the study were analyzed statistically by using one-way analysis of variance model in SAS version 9.2 Software packages. Based on significant differences of treatments, mean separation was done using Turkeys’ studentized (HSD) test. Mortality rate were corrected using Abbot (1925) formula. United State Environmental Protection Agency (USEPA) probit analysis version 1.5 was used for analyzing median lethal time and median lethal concentrations for each isolates at different concentrations (Finney, 1971). All recorded variables were analyzed according to the following statistical model:
where; Yij is any response, μ: is the general mean effect, Ti is the ith treatment effect and Eij is the experimental error.
RESULTS
Percentage mortality of worker termites: Percentage mortality of worker termites was calculated for the different concentrations of the four isolates and showed increasing mortality with increasing spore concentration. Cumulative mortality of worker termite over exposure period (one to seven days) was significantly different (p<0.05) for the fungi isolates (Fig. 1).
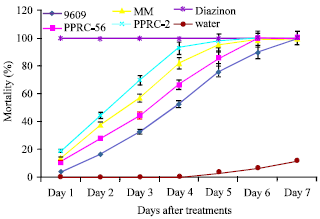 | |
| Fig. 1: | Cumulative Percent mortality of termite treated with M. anisopliae (MM and PPRC-2) and B. bassiana (9609 and PPRC-56) isolates |
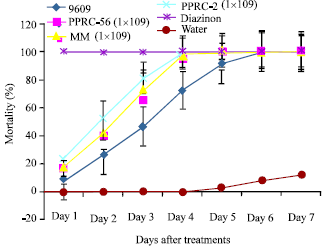 | |
| Fig. 2: | Percent Mortality of Macroterms treated with different fungal isolates at 1x109 conidia mL-1 (highest concentration) |
On the first day of exposure, maximum mortality (100%) was registered from standard check, while the untreated control had zero mortality. These were significantly different from all concentration of the isolates. Among the isolates, maximum mortality was observed from PPRC-2 isolates (19 to 100%) followed by MM isolates (14 to 100%) on day one up to day seven after treatment. Minimum mortality was registered from 9609 isolates (4 to 100%) followed by isolate PPRC-56 (11 to 100%) within the same period of exposure. Generally, as time progressed, cumulative mortality for all isolates increased attaining 100% mortality on the seventh day after application of the spore concentrations.
At the highest concentration, 1x109 conidial mL-1, all B. bassiana and M. anisopliae isolates gave complete reduction in population of worker termites resulting to 100% mortality after sixth day of exposure (Fig. 2). The initial data for pathogenicity of all the four isolates indicated that all of them were virulent, even one day after exposure causing significant mortality (up to 20%) when compared with the untreated control (zero mortality). The mortality percentage for all the isolates gradually increased and on sixth day after exposure all of the isolates gave 100% mortality similar to the standard control.
Mean percentage mortality of the worker termites due to the different isolates of the entomopathogenic fungi at different concentrations was significantly different over time (Table 2). After one day of exposure 100% and zero mortality was registered from standard and negative controls, respectively. Among the isolates, maximum and significant mortality (23.33%) was registered from PPRC-2 at highest concentration. However, this concentration was followed by and non-significantly different from PPRC-2 isolates (21.67%) at 1x108 conidia mL-1, PPRC-2 isolates (20.00%) at 1x107 conidia mL-1, PPRC-2 isolate (16.67%) at 1x106 conidia mL-1 and MM isolates (18.33%) at 1x109 conidia mL-1. Minimum mortality (1.67%) was recorded from 9609 isolates at lowest concentration followed by 9609 isolate (3.33%) at 1x106 conidia mL-1, (5%) at 1x107 conidia mL-1, (6.67% ) at 1x108conidia mL-1, (8.33%) at 1x109 conidia mL-1 and PPRC-56 isolate (5%) at 1x105 conidia mL-1 and these were on par with each others. This indicates that M. anisopliae isolate PPRC-2 at higher concentration is the most pathogenic fungi against termite followed with M. anisopliae isolate MM. On the second days after exposure, maximum mortality was also registered from standard check (100%). There was no mortality registered from untreated control (0%). From the isolates, maximum and significant mortality (53.33%) was registered from PPRC-2 at higher concentration followed by PPRC-2 (50%) at 1x108conidia mL-1, PPRC-2 (46.64%) at 1x107 conidia mL-1 and MM (46.67%) at 1x109 conidia mL-1 which were non-significantly different among each other. Significantly minimum mortality of 3.33% and 8.33% were recorded from 9609 isolates at 1x105 and 1x106 conidia mL-1, respectively which were also on par with the negative control.
On day three, highest and significant mortality (100%) was registered from standard check (Diazinon). No mortality was registered (0%) from control (Water). Highest and non-significant mortality of 81.67% was recorded from the isolates PPRC-2 at higher concentration followed by PPRC-2 at 1x108 conidial mL-1 (78.33%), MM isolate at 1x109 conidia mL-1 (73.33%) and PPRC-2 at 1x107 conidial mL-1 (71.67%) in that order. However, lower and significant mortality, 13.33 and 21.67%, were obtained from 9609 isolates at 1x105 and 1x106 conidial mL-1, respectively.
Similarly, on day four, standard check gave significantly higher worker termites’ mortality (100%) while no mortality (0%) was obtained from negative control.
| Table 2: | Mean percent mortality of termite treated with B. bassiana and M. anisopliae isolate at different concentrations over time |
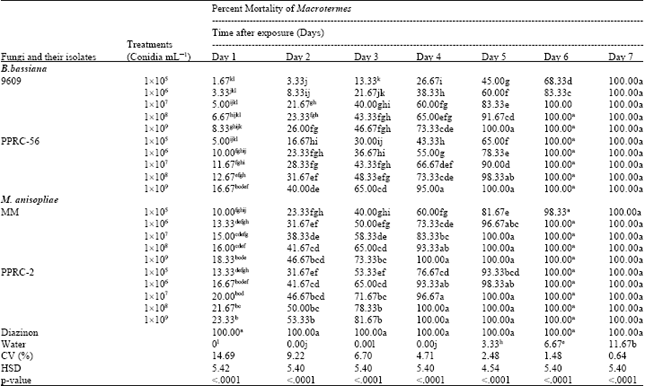 | |
| Means followed by the same letter within a column are not significantly different (Turkey’s Studentized range test, p<0.05) | |
Among the isolates, significant and higher mortality (100%) were recorded from the isolate PPRC-2 at 1x109conidia mL-1, 1x108conidia mL-1 and MM isolates at 1x109 conidia mL-1 followed by PPRC-2 (96.67%) at 1x107conidia mL-1, PPRC-56 (95.00%) at 1x109 conidia mL-1, PPRC-2 (93.33%) at 1x106 conidia mL-1 and MM isolate (93.33%) at 1x108 conidia mL-1. There were no significant differences among these concentrations of the respective isolates.
After fifth day of exposure, highest and significant mortality, 100%, of workers termite was registered from standard check; isolates PPRC-2 at 1x107 to 1x109 conidial mL-1, MM at 1x107 to 1x109 conidial mL-1, PPRC-56 at 1x109 conidial mL-1 and 9609 isolate at 1x109 conidial mL-1. Minimum significant mortality (3.33%) was recorded from untreated control. The mortality in the negative control treatment might be because of ageing. From the isolates, minimum significant mortality (45.00%) was registered from 9609 isolate at 1x105 conidial mL-1.
Six days after exposure, minimum and significantly lower percentage mortality was 6.67% from negative control treatment while the other treatments showed non-significant mortality among each other. From the isolates, 9609 isolates registered 68.33% at 1x105 conidial mL-1 and 83.33% at 1x106 conidial mL-1 where as isolate MM scored 98.33% mortality at 1x105 conidial mL-1. However, the rest isolates and standard checks, Diazinon were not significant different from each other with 100% mortality.
The trend on percent morality of worker termite gradually increased over exposure period and attained maximum mortality of 100% from sixth day on wards similar to standard check (Diazinon). In addition, it can be noticed that as concentration of the isolates increased, efficacy also increased on specific day after exposure.
Median lethal time (LT50) for fungal isolates: The median lethal time (LT50), time taken for the death of 50% of worker termite due to different isolates at different concentration (1x105 to 1x109conidia mL-1) were found to be different. From the isolates, M. anisopliae isolate PPRC-2 has shown the shortest median lethal time (7.74 days) at the highest concentration to observe 50% death of the target insect pests. On the other hand, median lethal time noted for the death of 50% of termite treated with B. bassiana isolates 9609 was the longest (8.80 days) at lowest concentration (1x105) (Table 3). Thus, the isolates of each entomopathogenic fungi were found to be promising in terms of the time taken as a management option against termite.
Median lethal concentration (LC50) for each fungal isolates: Results of the probit analysis obtained from applying known conidial suspensions of the two fungi isolates have shown that the LC50 for each isolate was different.
| Table 3: | Median lethal time (LT50) of B. bassiana (9609 and PPRC-56) and M. anisopliae (MM and PPRC-2) isolates against termite on day seven after treatment |
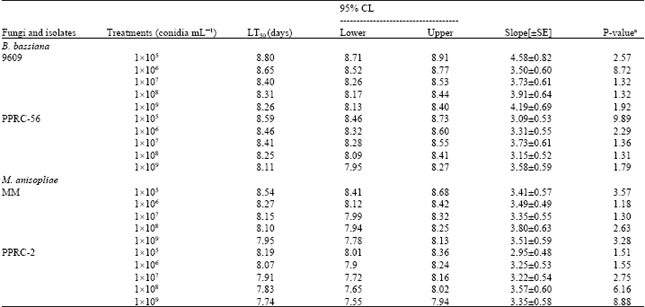 | |
| ais the single hypothesis one-sided p-value of the association between upper and lower limit (based on Fisher’s Exact Test) | |
| Table 4: | Median lethal concentration (LC50) of selected fungal isolates against termite, Macrotermes treated with aqueous conidial suspensions of different isolates of B.bassiana (9609 and PPRC-56) and M. anisopliae (MM and PPRC-2) on day seven |
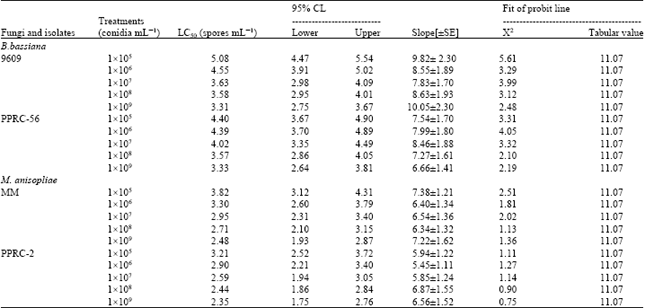 | |
| CL: Confidence limits, X2-Chi-Square | |
Among the two isolates of each M. anisopliae and B. bassiana tested; M. anisopliae isolate PPRC-2 had the least LC50 value (2.35x109 conidia mL-1) followed by PPRC-2 (2.44x108conidia mL-1) and MM (2.48x109conidia mL-1). While, the median lethal concentration to killed 50% of termite treated with B. bassiana isolates 9609 took the highest lethal concentration (5.08x105 conidia mL-1) followed by 9609 isolates (4.55x106 conidia mL-1), MM (4.40x105 conidia mL-1), MM (4.39x106 conidia mL-1) and MM (4.02x107conidia mL-1) (Table 4).
DISCUSSION
Spore concentrations of two species of entomopathogenic fungi (M. anosopliae (isolates MM and PPRC-2) and B. bassiana (9609 and PPRC-56) were applied on worker termite to determine fungi isolate(s) with high efficacy against Macrotermes under laboratory conditions. Both fungi isolates were found to be pathogenic to Macrotermes. However, there was a variation in their virulence against Macrotermes. The two isolates of B. bassiana were significantly less effective when compared with that of M. anosopliae isolates in terms of virulence. Among the isolates, PPRC-2, were found more virulent to the Macrotermes than the other isolates. Virulence due to M. anisopliae isolates on Macrotermes was not significantly different as compared to that of standard check after fifth days of the exposure period both causing 100% mortality. This indicates that from all the entomopathogenic fungi, M. anisopliae PPRC-2 isolate was the best entomopathogen for the control of Macrotermes. The next best entomopathogen was MM, followed by PPRC-56 and 9609 from B. bassiana in that order. These findings are in conformity with earlier reports (Singha et al., 2006; Ahmed et al., 2009; Dong et al., 2009; Pik-Kheng et al., 2009) who have shown a similar pattern of activity with isolates of these two entomopathogenic fungi against subterranean termite Coptotermes curvignathus. Both the fungal species were reported to produce an enzyme exoproteases with insecticidal activity.
The highest concentration (1x109conidia mL-1) of the fungal isolates M. anisopliae (PPRC-2 and MM) had nearly 100% mortality similar to the synthetic chemical (Diazinon) from day four on wards. On average, all the fungal isolates showed a high level of mortality, ranging from 25 to 75% (9609) and 45 to 95% (PPRC-56) for the isolate of B. bassian while 60 to 100% (MM) and 75 to 100% (PPRC-2) for M. anisopliae isolate when compared to 0.21% Diazinon with 100% mortality over time.
Currently the understanding of how effective the fungal isolates on Macrotermes was very limited due to less experiments, but many reports are available showing that the isolates cause a high mortality on other insects (Kannan et al., 2008). In this study it has been shown that all the fungal isolates are effective against termite and achieved appreciable mortality. An increase in the concentration of spores generally increased the mortality and generated a faster mortality.
After termites were infested with fungus spores, the mortality rate was lowest on the first two days and increased rapidly on the subsequent days. Percent mortality of termites depends on concentration of conidia and isolates of fungi. Termites that have been treated with conidia of M. anisopliae initiate to die within two days after inoculation. After six to seven days white mycelium had developed and green conidia were appeared around the insect cadavers. The reason for this is that fungus spores required the amount of time to thrive and sprout their mycelia into the termites (Krutmuanga and Mekchay, 2005; Brogden et al., 2005) opined that mortality increased after the second day of exposure because specific toxin, Destruxin, from the extracts penetrated swiftly into the termites’ hemocoel.
In this study, the calculated values of LT50 for the isolates suggest that, in general, the M. anisopliae isolates PPRC-2 (7.74 to 8.19 days) and MM (7.95 to 8.54 days) elicited mortality quickly than did the B. bassiana isolates 9609 (8.26 to 8.80 days) and PPRC-56 (8.11 to 8.59 days). This observation conforms to the earlier reports (Singha et al., 2006; Ahmed et al., 2009; Dong et al., 2009; Pik-Kheng, et al., 2009; Cherry et al., 2005). The median lethal time taken for fifty percent death of target insect (LT50) was more in the current result may be due to different factors such as difference in physiological combinations, methods of application of spore suspension and the amount used, the age of the insects used plus the condition where the experiments were conducted and other characteristics of entomopathogenic fungi. Also according to Todorova et al. (2002) and Alizadeh et al. (2007), lethal time (LT50) depends on the spore suspension and with the increase in spore suspension there is a decrease in time taken. Adane et al. (1996) reported the lowest times of 2.74 and 5.54 day for B. bassiana on bruchid and maize weevil, respectively.
The median lethal concentration (LC50) of different fungal isolates against termite, Macrotermes depend upon the fungal species or its isolate. Earlier studies confirmed that conidia of V. lecanii were highly pathogenic against aphids (Vu et al., 2007). The susceptibility of target insect to fungal infection is concentration dependent (Liu et al., 2002; Wright et al., 2005). Ansari et al. (2004) also reported that mortality of insect pests due entomopathogenic fungi depends on the concentration of conidial suspension, time of exposure and temperature. Entomopathogenic fungi M. anisopliae isolates had low LC50 values (3.21x105 to 3.82x105) compared to B. bassiana isolates (4.39x105 to 5.08x105) which is in line with the findings of Singha et al. (2011).
CONCLUSION
Based on this finding, we suggest that M. anisopliae and B. bassiana as a useful option for the management of Macrotermes spp. However, further investigations are strongly recommended to be carried out on the possibility of field application as well as finding other isolates of Entomopathogenic fungi that have potential as bio-termiticidal activities against such termite.
ACKNOWLEDGMENTS
We greatly acknowledge Ambo plant protection research center, Ethiopia for providing the test material, Entomopathogenic fungi (M. anisopliae and B. bassiana isolates). We are also indebted to thank Jimma University College of Agriculture and Veterinary Medicine for providing laboratory space and facilities as well as financial support to conduct the study.
REFERENCES
- Abbott, W.S., 1925. A method of computing the effectiveness of an insecticide. J. Econ. Entomol., 18: 265-267.
CrossRefDirect Link - Adane, K., D. Moore and S.A. Archer, 1996. Preliminary studies on the use of Beauveria bassiana to control Sitophilus zeamais (Coleoptera: Curculionidae) in the laboratory. J. Stored Prod. Res., 32: 105-113.
CrossRefDirect Link - Ahmed, S., M.R. Ashraf, A. Hussain and M.A. Riaz, 2009. Pathogenicity of isolates of Metarhizium anisopliae from Gujranwala (Pakistan) against Coptotermes heimi (wasmann) (Isoptera: Rhinotermitidae). Int. J. Agric. Biol., 11: 707-711.
Direct Link - Alizadeh, A., A.K. Pakdel, K.H. Talebi-Jahromi and M.A. Samih, 2007. Effect of some Beauveria bassiana (Bals.) Viull. isolates on common pistachio psylla Agonoscena pistaciae Burck. and Laut. Int. J. Agri. Biol., 9: 76-79.
Direct Link - Abera, A., F. Lemessa and D. Muleta, 2011. The antifungal activity of some medicinal plants against coffee berry disease caused by Colletotrichum kahawae. Int. J. Agric. Res., 6: 268-279.
CrossRefDirect Link - Ansari, M.A., S. Vestergaard, L. Tirry and M. Moensa, 2004. Selection of a highly virulent fungal isolates, Metarhizium anisopliae CLO 53, for controlling Hoplia philanthus. J. Invertebrate Pathol., 85: 89-96.
CrossRefDirect Link - Bahiense, T.C., E.K.K. Fernandes and V.R.E.P. Bittencourt, 2006. Compatibility of the fungus M. anisopliae and deltamethrin to control a resistant isolate of Boophilus microplus tick. Vet. Parasitol., 141: 319-324.
CrossRef - Bittencourt, V.R., 2000. Trials to control South American ticks with entomopathogenic fungi. Ann. Nat. Acad. Sci., 916: 555-558.
PubMedDirect Link - Brogden, K.A., J.M. Guthmiller, M. Salzet and M. Zasloff, 2005. The Nervous System and Innate Immunity: The Neuropeptide Connection. Nat. Immunol., 6: 558-564.
CrossRef - Cherry, A.J., P. Abalo and K. Hell, 2005. A laboratory assessment of the potential of different strains of the entomopathogenic fungi Beauveria bassiana (Balsamo) Vuillemin and Metarhizium anisopliae (Metschnikoff) to control Callosobruchus maculatus (F.) (Coleoptera: Bruchidae) in stored cowpea. J. Stored Prod. Res., 41: 295-309.
CrossRefDirect Link - Singha, D., B. Singha and B.K. Dutta, 2011. Potential of Metarhizium anisopliae and Beauveria bassiana in the control of tea termite Microtermes obesi Holmgren in vitro and under field conditions. J. Pest Sci., 84: 69-75.
CrossRef - Dong, C., J. Zhang, H. Huang, W. Chen and Y. Hu, 2009. Pathogenicity of new China variety of Metarhizium anisopliae (M. anisopliae var. Dcjhyium) to subterranean termite Odontotermes formosanus. Microbiol. Res., 164: 27-35.
CrossRef - Finney, D.J., 1971. Probit Analysis. 3rd Edn., Cambridge University Press, London, UK., pp: 76-80.
CrossRefDirect Link - Gitonga, W., N.K. Maniania, J. Eilenberg and G.R.S. Ocheil, 1995. Pathogenecity of Indigenous Enthomopathogenic Fungi Metarhizium anispliae and Beauveria bassiania to Macrotermes michaelseni in Kenya. Proceedings of the 2nd Regional Workshop on Termite Research and Control, March 7-9, 1995, Nairobi, Kenya, pp: 36-44.
- Hoe, P.K., C.F.J. Bong, K. Jugah and A. Rajan, 2009. Evaluation of Metarhizium anisopliae var. anisopliae (Deuteromycotina: Hyphomycete) isolates and their effects on subterranean termite Coptotermes curvignathus (Isoptera: Rhinotermitidae). Am. J. Agric. Biol. Sci., 4: 289-297.
CrossRefDirect Link - Jaramillo, J. and C. Borgemeister, 2006. New bioassay method to assess the pathogenicity of Colombian strains of Metarhizium anisopliae (Metsch.) Sorokin and Paecilomyces sp. (Deuteromycotina: Hyphomycetes) against the subterranean burrower bug Cyrtomenus bergi Froeschner (Hemiptera: Cydnidae). J. Invertebr. Path., 91: 57-60.
CrossRefPubMed - Julia, G. and H. Lina, 2010. The potential of entomopathogenic fungal isolates as an environmentally friendly management option against Acanthoscelides obtectus. Student Degree Project for Environmental Management, Swedish University of Agricultural Sciences and Addis Ababa University, Addis Ababa, Ethiopia, Pages: 26.
- Kaaya, G.P., 2003. Prospects for innovative methods of tick control in Africa. Insect Sci. Application, 23: 59-67.
CrossRef - Liu, H., M. Skinner, L.P. Bruce and M. Brownbridge, 2002. Pathogencity of Beauveria bassiana, Metarhizium anisopliae (Deuteromycotina: Hyphomycetes) and other Entomopathogenic Fungi against Lygus lineolaris (Hemiptera: Miridae). J. Econ. Entomol., 95: 675-681.
CrossRef - Logan, J.W.M., R.H. Cowie and T.G. Wood, 1990. Termite (Isoptera) control in agriculture and forestry by non-chemical methods: A review. Bull. Entomol. Res., 80: 309-330.
CrossRefDirect Link - Maniania, N.K., S. Sithanantham, S. Ekesi, K. Ampong-Nyarko, J. Baumgartner, B. Lohr and C.M. Matoka, 2003. A field trial of the entomopathogenic fungus Metarhizium anisopliae for control of onion thrips, Thrips tabaci. Crop Prot., 22: 553-559.
CrossRef - Milner, R.J., 2003. Application of biological control agents in mound building termites (Isoptera: Termitidae): Experiences with Metarhizium in Australia. Sociobiology, 41: 419-428.
Direct Link - Milner, R.J., J.A. Staples and G.G. Lutton, 1998. The selection of an isolate of the Hyphomycete fungus, Metarhizium anisopliae, for control of termites in Australia. Biol. Control, 11: 240-247.
CrossRef - Nielsen, C., H. Skovgard and T. Steenberg, 2004. Effect of Metarhizium anisopliae (Deuteromycotina: Hyphomycetes) on survival and reproduction of the filth fly parasitoid, Spalangia cameroni (Hymenoptera: Pteromalidae). Environ. Entomol., 34: 133-139.
CrossRefDirect Link - Nyeko, P., L.S. Glohole, N.K. Maniania, H. Agaba and B.M. Sekamate, 2010. Evaluation of Metarhizium ansophiliae for integrated management of termite on maize and Grevillea rebusta in Uganda and Kenya. Proceedings of the 2nd RUFORUM Biennial Meeting, September 20-24, 2010, Entebbe, Uganda, pp: 333-336.
- Osipitan, A.A. and A.E. Oseyemi, 2012. Evaluation of the bio-insecticidal potential of some tropical plant extracts against termite (Termitidae:Isoptera) in Ogun State, Nigeria. J. Entomol., 9: 257-265.
CrossRefDirect Link - Samb, T., A.B. Ndiaye and K. Diarra, 2011. Biodiversity of termites in relation to human activity: Impact on the environment in Matam (Senegal). Res. J. Pharm. Biol. Chem. Sci., 2: 313-323.
Direct Link - Samson, P.R., R.J. Milner, E.D. Sander and G.K. Bullard, 2005. Effect of fungicides and insecticides applied during planting of sugarcane on viability of M. anisopliae and its efficacy against white grubs. Bio Control, 50: 151-163.
CrossRef - Scholte, E.J., B.C.J. Knols and W. Takken, 2006. Infection of the malaria mosquito Anopheles gambiae with the entomopathogenic fungus Metarhizium anisopliae reduces blood feeding and fecundity. J. Invertebrate Pathol., 91: 43-49.
CrossRefDirect Link - Sharma, N., K.P. Sharma, R.K. Gaur and V.K. Gupta, 2011. Role of chitinase in plant defense. Asian J. Biochem., 6: 29-37.
CrossRefDirect Link - Singha, D., B. Singha and B.K. Dutta, 2006. Virulence of isolates of Metarhizium anisopliae (Metschnikoff) Sorokin and Beauveria bassiana (Ballsamo) Vuillemin on tea termite Microcerotermes beesoni Snyder Barak valley of Assam. Indian J. Entomol., 68: 66-70.
Direct Link - Suliman, E.A. and Y.O. Mohammed, 2012. The activity of Aspergillus terreus as entomopathogenic fungi on different stages of Hyalomma anatolicum anatolicum under experimental conditions. J. Entomol., 9: 343-351.
CrossRefDirect Link - Sun, J., J.R. Fuxa and G. Henderson, 2003. Effects of virulence, sporulation and temperature on Metarhizium anisopliae and Beauveria bassiana laboratory transmission in Coptotermes formosanus. J. Invert. Pathol., 84: 38-46.
Direct Link - Todorova, S.I., C. Cloutier, J.C. Cote and D. Coderre, 2002. Pathogenicity of six isolates of Beauveria bassiana (Balsamo) Vuillemin (Deuteromycotina, Hyphomycetes) to Perillus bioculatus (F) (Hem., Pentatomidae). J. Applied Entomol., 126: 182-185.
CrossRef - Vu, V.H., I.S. Hong and K. Kim, 2007. Selection of entomopathogenic fungi for aphid control. J. Biosci. Bioeng., 104: 498-505.
CrossRef - Wang, C. and J.E. Powell, 2004. Cellulose bait improves the effectiveness of termites (Isoptera: Rhinotermitidae). Biol. Control, 30: 523-529.
CrossRef - Wright, M.S., A.K. Raina and A.R. Lax, 2005. A strain of the fungus Metarhizium anisopliae for controlling subterranean termites. J. Econ. Entomol., 98: 1451-1458.
PubMedDirect Link - Zimmermann, G., 1993. The entomopathogenic fungus Metarhizium anisopliae and its potential as a biocontrol agent. Pest. Sci., 37: 375-379.
CrossRef