
Akram Nasiri
Department of Biology, Faculty of Science, University of Isfahan, Isfahan, Iran
Hojjatollah Saeidi
Department of Biology, Faculty of Science, University of Isfahan, Isfahan, Iran
Mohamad Reza Rahiminejad
Department of Biology, Faculty of Science, University of Isfahan, Isfahan, Iran
Asian Journal of Plant Sciences
Year: 2012 | Volume: 11 | Issue: 2 | Page No.: 62-69







ABSTRACT
Cynodon dactylon is widely distributed in Iran in diverse habitats. In this study, morphological variation of 46 populations of this species was evaluated based on 52 qualitative and quantitative characters to reveal the true taxonomic situation of the species. High morphological variation with no significant correlation with geographical and ecological conditions was observed. Among the morphological characters, presence of hair on leaf and sheath surfaces and spike color had major role on groupings in dendrogram. Based on the results of this study, presence of pilose hairs on leaf surfaces and outer surface of leaf sheath were shown to be stable diagnostic characters in detecting infraspecific taxa and accordingly, the species C. dactylon present in Iran two varieties: var. dactylon and var. villosus Regel. A morphological description for this species and taxonomic key of the varieties are provided.
PDF Abstract XML References Citation
Received: January 20, 2012;
Accepted: March 05, 2012;
Published: May 14, 2012
How to cite this article
Akram Nasiri, Hojjatollah Saeidi and Mohamad Reza Rahiminejad, 2012. Morphological Variation and Taxonomic Conclusion of Cynodon dactylon (L.) Pers. In Iran. Asian Journal of Plant Sciences, 11: 62-69.
DOI: 10.3923/ajps.2012.62.69
URL: https://scialert.net/abstract/?doi=ajps.2012.62.69
DOI: 10.3923/ajps.2012.62.69
URL: https://scialert.net/abstract/?doi=ajps.2012.62.69
INTRODUCTION
Cynodon dactylon (L.) Pers. (common bermudagrass) is a perennial herb belonging to the tribe Chlorideae (Poaceae). Its role as turf grass, forage and its function in soil stabilization and conservation (Roudsari and Pishdar, 2007), protecting the soil of erosion and also its nature as weed (Oad et al., 2007; Mahanta et al., 2007) and medicinal plants (Nagori and Solanki, 2011; Chaudhary et al., 2001; Garg and Paliwal, 2011; Mahesh and Brahatheeswaran, 2007) made this species as an economically importance species (Bor, 1968; Assefa et al., 1999). Today, it is widely used as warm-season turf and forage species in the temperate and tropical regions (Li and Qu, 2004). The populations of this species harbor many useful physiological traits such as tolerance to high temperature and drought (Bethel, 2005).
This species was validated by Linnaeus (1753) as Panicum dactylon L. Persoon (1805) transferred it to the monotypic genus Cynodon Rich. as Cynodon dactylon (L.) Pers.. Morphological complexities resulted in different infraspecific classifications in the species. In taxonomic revision of the Cynodon genus published by Harlan et al. (1970), this species comprises six varieties include var. dactylon, var. afghanicum Harlan et de Wet, var. aridus Harlan et de Wet, var. coursii (A. camus) Harlan et de Wet, var. elegans Rendle and var. polevansii (Stent) Harlan et de Wet. Meanwhile, Cynodon dactylon var. dactylon is the most widely distributed and genetically variable variety in this species. Rozhevits and Shishkin (1934), Tutin et al. (1980) and Nasir and Ali (1982) recognized no subspecies or variety for it. Two varieties: var. dactylon and var. villosus Regel were recognized by Bor (1968) based on absence/presence of villous hairs on racemes and by Davis (1985) based on absence/presence of pilose hairs on leaf blades and/or leaf surfaces. Wu et al. (2006) subdivided this species into two varieties dactylon and biflorus Merino. based on having one or two bisexual floret in spikelet.
Harlan and de Wet (1969) suggested that the species is originated and diversified somewhere from West Pakistan to Turkey (i.e., Turkey, Iran, Afghanistan and the West part of Pakistan) and distributed in all countries and islands between about lat 45°N and lat 45°S and penetrates to approximately lat 53°N in Europe. Information on the genetic and cytological diversity of C. dactylon in its center of diversity is meager.
The taxonomic status and morphological diversity of this species in Iran is unclear. This study aimed to revise the taxonomic status of C. dactylon in Iran and to evaluate the morphological variation of this species and find out the taxonomic characters separating possible taxa belonging to this species in Iran.
MATERIALS AND METHODS
A total of 460 individual plants belonging to 46 populations of C. dactylon were specifically collected from various regions of Iran for this study (Table 1). The specimens were collected from regions with different ecological conditions.
| Table 1: | Collection site of 46 Iranian populations of Cynodon dactylon analysed in this study |
 | |
| * C: Center, W: West, N: North, NW: Northwest, S: South, E: east, SE: Southeast and SW :Southwest. | |
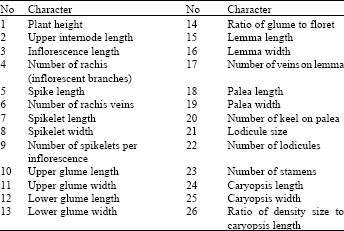
Taxonomic identifications were made based on Bor (1970). Variability of 59 qualitative and 26 quantitative morphological characters were evaluated among the specimens. Twenty-six out of 59 qualitative characters showed variability among individuals and were used for further analyses together with 26 quantitative characters (Table 2, 3). In multistate qualitative characters, each state was considered as one independent character and converted to a binary (absent/present) character. The quantitative characters were standardized using formulae:
where, Xs is the standardized data, x is the raw data, min and max are the minimum and maximum amount of the data and n is number of observations. Data were analysed using NTSYS-pc 2.02 software (Rohlf, 1998).
Different similarity coefficients were used to calculate morphological similarities. To test the goodness of fit of a cluster analysis to our data, the cophenetic value matrix from each tree matrix was computed, which was then compared with its corresponding similarity matrix (matrix comparison) to compute the matrix correlation value (r value). The simple matching coefficient (Sneath and Sokal, 1973) based similarity matrix showed the highest r value and was used to generate dendrograms.
| Table 2: | List of evaluated variable qualitative morphological characters used for diversity analysis in Cynodon dactylon in Iran |
 | |
| Table 3: | List of evaluated quantitative morphological characters in species Cynodon dactylon |
 | |
Dendrograms were generated after cluster analysis with the unweighted pair group method using arithmetic averaging (UPGMA).
RESULTS AND DISCUSSION
Morphological diversity: Analysis of morphological data showed high variability among the populations studied. In cluster analysis, the reproductive characters had higher influence on groupings of populations in dendrogram. Status of leaf surface and abaxial sheath surface hairs were major factors separating the dendrogram main branches.
In dendrogram generated based on the qualitative characters (Fig. 1), populations were divided in two major groups with 61% similarity value (groups A and B in Fig. 1). These two groups were different mainly in status of hairs on both surfaces of leaf and outer surface of sheath and spike color. The group A comprises those populations with pale-green spikes and without pilosus hairs on abaxial leaf and sheath surfaces. Within this group, in population 45 (subgroup A2), some individuals had pilosus hairs on abaxial leaf and sheath surfaces and was separated from others (subgroup A1). Further subdivisions within these groups were made by present or absent of tomentose hairs on both sides of leaf and sheaths (subgroups A1a and A1b in Fig. 1).
Group B was subdivided into two groups based on present or absent of pilosus hairs on both sides of leaf and outer surface of sheath and violet spots on leaf surface. Populations located in subgroup B1 lacking pilosus hairs on one of the above surfaces and had violet spots, however, the populations in subgroup B2 had both characters together. Populations in group B1 were divided into two subgroups based on present of sparse hairs on abaxial leaf surface and absent of it on outer sheath surface (subgroups B1a and B1b). Group B2 were divided into subgroups (B2a and B2b).
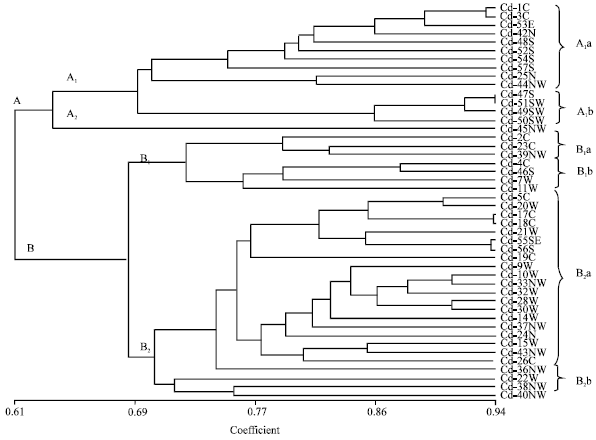 | |
| Fig. 1: | UPGMA dendrogram based on 26 variable qualitative characters analysed among 46 population of Cynodon dactylon collected from Iran using simple matching coefficient. Population numbers are shown with geographic origin (N, NW, W, S, SE, SW, E and C, Table 1) |
Subgroup B2b comprises those populations with short pilose hairs on abaxial leaf surface and dense pilose hairs on outer sheath surface.
Hair presence on leaf and sheath surfaces were important characters analysed in many previous studies (Boissier, 1884; Rozhevits and Shishkinm, 1934; Bor, 1970; Davis, 1985). Rozhevits and Shishkin (1934) reported two forms glabrum Roshev. and villosum Rgl. within this species based on presence/absence of hairs on leaf surface. Presence or absence of hairs on leaf and sheath were used by Davis (1985) as diagnostic characters for distinguish var. dactylon and var. villosus Regel within this species.
Based on the results of this study there were no correlation between ecological conditions and presence and density of hairs. This character was diagnostic for distinguishing infraspecific taxa. Despite being highly variable, other qualitative characters showed no variation related to the infraspecific taxa.
Variability of quantitative characters was considerably lower than that of the quantitative ones. In dendrogram generated based on quantitative characters (Fig. 2), populations were divided into two groups at 53% similarity. Difference in the width of the lower glume had major role in this grouping. Group A was subdivided into two subgroups based on Inflorescent length (subgroups A1 and A2). Also stem length had major role in subgroup formations within group A1 (subgroups A1a and A1b). The group B was subdivided into subgroups based on differences in the number of nerves on the rachis, length and width of upper and lower glumes. Dendrogram generated based on a combination of qualitative and quantitative characters (Fig. 3) showed high similarity in topology and branching with that constructed based on qualitative characters only, indicating that the diversity presented in this species is mainly due to the variation in qualitative characters.
Regarding high apomixy and outbreeding in this species, it can be suggested that different genotypes of this species colonized first in different regions and then, outbreeding distributed this genetic diversity all over the different regions. This high diversity then was established by apomixy.
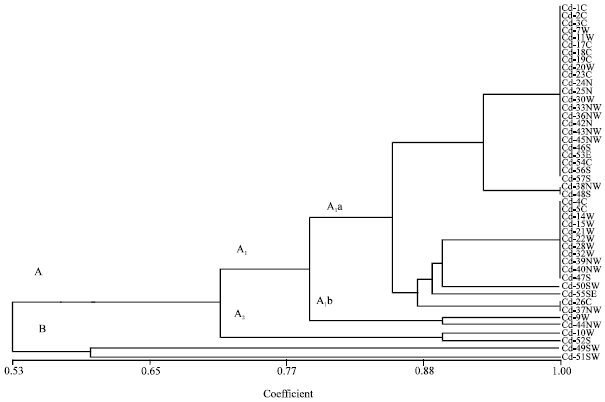 | |
| Fig. 2: | UPGMA dendrogram based on 26 standardized quantitative characters examined among 46 population of Cynodon dactylon collected from Iran using simple matching coefficient. Population names are shown with geographic origin (N, NW, W, S, SE, SW, E and C, Table 1) |
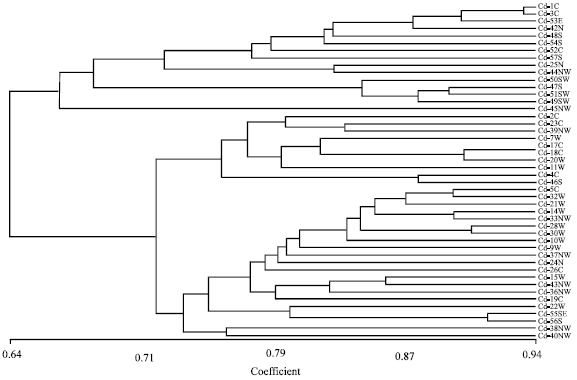 | |
| Fig. 3: | UPGMA dendrogram based on 26 variable qualitative and 26 standardized quantitative characters examined among 46 population of Cynodon dactylon collected from Iran using simple matching coefficient. Population numbers are shown with geographic origin (N, NW, W, S, SE, E, SW and C, Table 1) |
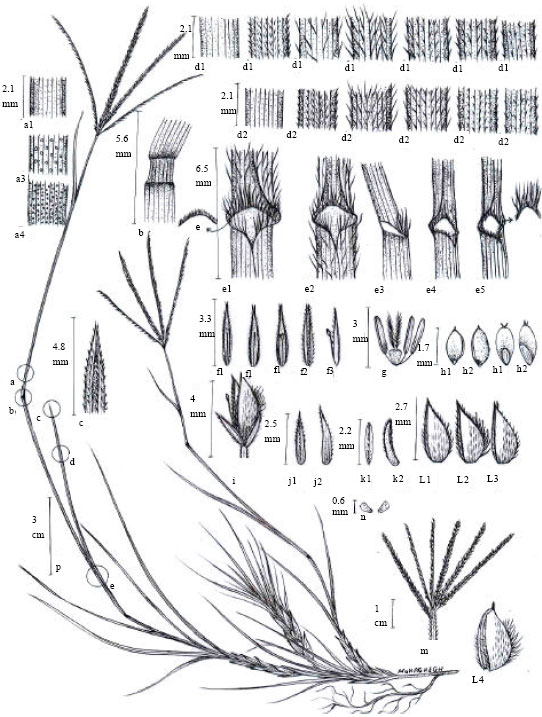 | |
| Fig. 4: | Cynodon dactylon (L.) Pers., A: Plant habit, a1-4: Stem, b: node, c: Leaf marginal ornamentations, d1: Leaf upper surface, d2: Leaf lower surface, e: Ligule, e1-5: Leaf sheath, f1-3: Palea, g: Anthers and pistil (flower), h1-2: Seed, i: Spikelet, j1-2: Upper glume, k1-2: Lower glume, l1-4: Lemma, m: Inflorescence, n: Lodicule |
Taxonomy:
| • | Cynodon dactylon (L.) Pers., Syn. Pl. 1: 85 (1805) |
| • | Syn: Panicum dactylon L., Sp. Pl. 58 (1753) |
Herbaceous, perennial, rhizomatous and with creeping stem. Rhizomes branched, scaled, glabrous, light pink to yellowish. Creeping stem long and rooted at nods. Flowering stems many, erect, ascendant, 7-49.5 cm, usually glabrous, rarely hairy, ended to a set of spikes 2-8 cm long (Fig. 4 and 5). Leaves linear – lanceolate, flat, acute, green and scabrous at margins. Both surfaces of leaf and outer surface of sheat glabrous or with short or long hairs. Ligule a fringe of white hairs.
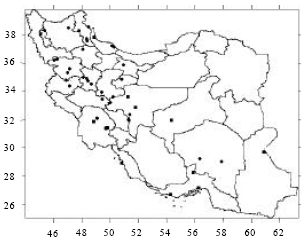 | |
| Fig. 5: | Collection sites of population of species Cynodon dactylon (L.) Pers. in Iran. var. villosus Regel (•) and var. dactylon (■) analysed in this study |
Inflorescent raceme-like panicle, pilosus at base and with 3-6 rarely to 9 spikes. Spikes glabrous, elongated, green to pale green, or green with purple spots, yellowish at maturity. Rachis scabrid and ribbed. Spikelet ovate, 2-3 mm, usually purplish, stalked, awnless, oriented at one side of rachis, with one fertile and one aborted florets. Glumes unequal and shorter than florets. Upper glum longer than the lower, 1.1-2.5x0.6-0.8 mm, acute, awnless, keeled, scabrous at abaxial surface of the keel, with a green rib at the keel. Lower glume membranous, 1.1-2.2x0.4-0.8 mm, keeled to falciform, glabrous, acuminate, awnless, scabrous at keel. Lemma ovate to oblong, acute to acuminate, keeled, green to pale-green, with purple spots, glabrous, with three nerves, 1.9-2.7x0.7-1.1 mm. palea linear, elongated, acuminate, with tow keels, usually with short and dense hairs at keels, 1.7-2.8x0.3-0.5 mm. Lodicules 2, membraneous, clavate. Stamens 3, yellowish to purple, with big anthers. Grain oblong, glabrous.
| • | Flowering time: May-June |
| • | Distribution: cosmopolitan |
| • | Distribution in Iran: this species distributed in different regions of Iran with a vast range of environmental conditions, from sea side to above 2700 m |
Based on the results of this study, there are tow varieties of this species in Iran that can be distinguished by taxonomic key as below:
| • | Plant without hair ……………var. dactylon |
| • | Plant with hairs at least at one sides of leaf surfaces or outer surface of sheath …………..var. villosus |
REFERENCES
- Assefa, S., C.M. Taliaferro, M.P. Anderson, B.G. de los Reyes and R.M. Edwards, 1999. Diversity among Cynodon accessions and taxa based on DNA amplification fingerprinting. Genome, 42: 465-474.
PubMed - Chaudhary, M.I., A.S. Mumtaz and M.A. Khan, 2001. Leaf epidermal anatomy of medicinal grasses of Islamabad, Attock and Mirpur (Azad Kashmir). Pak. J. Biol. Sci., 4: 1466-1469.
CrossRefDirect Link - Garg, V.K. and S.K. Paliwal, 2011. Anti-inflammatory activity of aqueous extract of Cynodon dactylon. Int. J. Pharmacol., 7: 370-375.
CrossRefDirect Link - Harlan, J.L. and J.M.J. de Wet, 1969. Sources of variation in Cynodon dactylon (L.) Pers. Crop Sci., 9: 774-778.
CrossRefDirect Link - Li, L. and R. Qu, 2004. Development of highly regenerable callus lines and biolistic transformation of turf-type common bermudagrass [Cynodon dactylon (L.) Pers.]. Plant Cell Rep., 22: 403-407.
CrossRefDirect Link - Mahanta, J.J., M. Chutia and T.C. Sarma, 2007. Study on weed flora and their influence on patchouli (Pogostemon cablin benth.) oil and patchoulol. J. Plant Sci., 2: 96-101.
CrossRefDirect Link - Mahesh, N. and D. Brahatheeswaran, 2007. Anti-hyperglycemic activities of aqueous and ethanolic extracts Cynodon dactylon (Linn) streptozotocin-induced diabetic rats. Asian J. Biochem., 2: 66-72.
CrossRefDirect Link - Nagori, B.P. and R. Solanki, 2011. Cynodon dactylon (L.) Pers.: A valuable medicinal plant. Res. J. Med. Plant, 5: 508-514.
CrossRefDirect Link - Oad, F.C., M.H. Siddiqui, U.A. Buriro and G.S. Solangi, 2007. Weed management practices in cotton crop. Asian J. Plant Sci., 6: 344-348.
CrossRefDirect Link - Roudsari, O.N. and H. Pishdar, 2007. Evaluation of Composted Sewage Sludge (CSS) as a soil amendment for bermudagrass growth. Pak. J. Biol. Sci., 10: 1371-1379.
CrossRefPubMedDirect Link - Rohlf, F., 1998. NTSYS-pc: Numerical Taxonomy and Multivariate Analysis System. Version 2.2, Exeter Publications Setauket, New York.
Direct Link