
A. H. Gorji
Department of Plant Breeding, Faculty of Agriculture, Broujerd Branch,Islamic Azad University, Broujerd, Iran
Z. Zonoori
Department of Plant Breeding, Faculty of Agriculture, Broujerd Branch,Islamic Azad University, Broujerd, Iran
M. Zolnoori
Department of Plant Breeding, Faculty of Agriculture, Broujerd Branch,Islamic Azad University, Broujerd, Iran
A. Jamasbi
Department of Plant Breeding, Faculty of Agriculture, Broujerd Branch,Islamic Azad University, Broujerd, Iran
Asian Journal of Plant Sciences
Year: 2011 | Volume: 10 | Issue: 3 | Page No.: 220-226







ABSTRACT
The objective of this study was to evaluate the inheritance of antioxidant activity of triticale under drought stress. For this study, the F1 seeds of a 7x7 half diallel along with their parents were grown in greenhouse in well watered and drought conditions. For this purpose, seven selected lines from breeding programs conducted previously were used as plant entries of the research. Yield and yield components were taken up to determine the best lines to be candidate for future varieties. After exposure of plants to drought stress, Catalase (CAT), the activity of Ascorbate Peroxidase (APX) and Superoxide Dismutase (SOD) enzymes were measured. Presence of over dominance and greater importance of dominance effects in control of traits was observed. All of traits had very high broad sense heritability. In stress condition among the traits APX had high narrow sense heritability (Hn), followed by moderate Hn for CAT and low Hn for SOD and in normal condition APX had highest Hn, followed by SOD with moderate Hn and CAT with low Hn. Regarding to the significant correlations between drought tolerance and antioxidant activity in the literature and considering the highest heritability of antioxidant enzymes in this experiment, it can be inferred that they can be considered as good yardstick for selecting resistance to drought in triticale.
PDF Abstract XML References Citation
Received: April 14, 2011;
Accepted: May 31, 2011;
Published: July 28, 2011
How to cite this article
A. H. Gorji, Z. Zonoori, M. Zolnoori and A. Jamasbi, 2011. Inheritance of Antioxidant Activity of Triticale under Drought Stress. Asian Journal of Plant Sciences, 10: 220-226.
DOI: 10.3923/ajps.2011.220.226
URL: https://scialert.net/abstract/?doi=ajps.2011.220.226
DOI: 10.3923/ajps.2011.220.226
URL: https://scialert.net/abstract/?doi=ajps.2011.220.226
INTRODUCTION
Adequate water and nutrient supply are important factors affecting optimal plant growth and successful crop production. Water stress is one of the severe limitations of crop growth especially in arid and semiarid regions of the world as it has a vital role in plant growth and development at all growth stages (Ahmad et al., 2009). Plants respond to diverse environmental signals in order to survive stresses such as drought (Pastori and Foyer, 2002). Drought leads to oxidative stress in plants due to the production of Reactive Oxygen Species (ROS) such as the super oxide radical, hydrogen peroxide and hydroxyl radical. Oxidative stress is one of the major limiting factors in plant productivity. Reactive Oxygen Species (ROS) generated during metabolic processes damage cellular functions and consequently lead to disease, senescence and cell death. Plants have evolved an efficient defense system by which the ROS is scavenged by antioxidant enzymes such as Superoxide Dismutase (SOD), Calatase (CAT), Peroxidase (POX), Polyphenoloxidase (PPO) and Glutathione Reductase (GR) (Joseph and Jini, 2010). Drought stress invariably leads to oxidative stress in the plant cell due to higher leakage of electrons towards O2 during photosynthetic and respiratory processes leading to enhancement in activated oxygen species generation (Stepien and Klobus, 2005). Photosynthesis is particularly sensitive to water deficit because the stomatal closure to conserve water, depletes intercellular CO2. This process reduces the availability of CO2 for photosynthesis which can lead to the formation of Reactive Oxygen Species (ROS) from the misdirecting of electrons in the photosystem (Reddy et al., 2004). Antioxidants are compounds that inhibit or delay the oxidation of other molecules by inhibiting the initiation or propagation of oxidizing chain reactions (Klein and Kurilich, 2000). Antioxidants function by scavenging free radicals via donation of an electron or a hydrogen atom, or by deactivation of prooxidant metal ions and singlet oxygen (Shahidi, 2002). Morello et al. (2002) stated that the primary role of antioxidants is to prevent degradation induced by free radical reactions. They noted that antioxidants function by hydrogen abstraction and metal ion assisted electron transfer. The antioxidant donates hydrogen atoms to the free radicals, thus inhibiting the propagation of the autocatalytic chain reaction. Drought-induced process of stomatal closure increases the oxidative load on the plant tissue. This causing imbalance in biochemical phatways and consequently formation of Reactive Oxygen Species (ROS), such as super oxideradicals, hydrogen peroxide, singlet oxygen and hydroxyl radicals (Luna et al., 2005). Stress resistance in plants is a complex character that depends on many genes and thus is determined by the interactions of many morphological, physiological and biochemical processes. Antioxidative system in plants has been composed of several enzymatic and non-enzymatic components that are being active in a differential manner in response to drought stress in different plants (Gholizadeh, 2010). There are different arrays of mechanisms that plant breeders use to study the adaptive response of different genotypes and as criteria for selection under unfavorable conditions (Hefny and Abdel-Kader, 2007). Usually, oxidative system is controlled/balanced by antioxidative system during the normal growth and developments in plant system. Although, plants commonly activate their total antioxidation machinery in response to drought stress situations but it has been well shown that different plants exhibit different responses to stress in terms of various redox enzymes activities. Habibi et al. (2004) and also Saleh and Plieth (2009) in theirs studies have been shown that drought stress in plants induces the oxidative burst that is obviously followed by the activation of their antioxidative system (Gholizadeh, 2010). Oxidative stress is also a factor in abiotic and biotic stress phenomena that occurs when there is a serious imbalance between the production of Reactive Oxygen Species (ROS) and antioxidant defense. ROS have been considered mainly as dangerous molecules and their concentrations must be maintained as low as possible (Amirjani, 2010). Chelikani et al. (2004) indicated that Catalase is a common enzyme found in nearly all living organisms that are exposed to oxygen, where it functions to catalyze the decomposition of hydrogen peroxide to water and oxygen. It is a tetramer of four polypeptide chains, each over 500 amino acids long (Joseph and Jini, 2010). Molecular genetic maps have been developed for major crop plants, including rice, wheat, maize, barley, sorghum and potato, which make it possible for scientists to tag desirable traits using known DNA landmarks. Molecular genetic markers allow breeders to track genetic loci controlling stress resistance without having to measure the phenotype, thus reducing the need for extensive field-testing over time and space. Moreover, gene pyramiding or introgression can be done more precisely using molecular tags. Together, molecular genetic markers offer a new strategy known as marker assisted selection. Another molecular strategy which depends on gene cloning and plant transformation technology, is genetic engineering of selected genes into elite breeding lines. What makes a particular goal attainable or unattainable in genetic engineering experiments is the availability of the following three inputs: (1) the gene of interest, (2) an effective technique for transferring the desired gene from one species to another and (3) promoter sequences for regulated expression of that gene. Amongst these, the first is considered a rate-limiting factor (Joseph and Jini, 2010). Higher plants have active oxygen scavenging systems consisting of several antioxidant enzymes, such as Superoxide Dismutase (SOD), Ascorbate Peroxidase (APX), Catalase (CAT) and Glutathione Reductase (GR) and some low molecules of non-enzyme antioxidants (Grene, 2002). These cytotoxic oxygen species are highly reactive and in the absence of any protective mechanism they can seriously disrupt normal metabolism through oxidative damage resulting in lipid peroxidation and consequently membrane injury, protein degradation, consequently membrane injury, protein degradation, enzyme inactivation, pigment bleaching and disruption of DNA strands (Pan et al., 2006; Quiles and Lopez, 2004). Antioxidant enzymes can be measured easily and are nondestructive to whole plant. So they potentially can be considered as selection criteria if they also have high heritability. The objective of this study was to evaluate the inheritance of heritability under drought stress.
MATERIALS AND METHODS
Seven triticale lines (Table 1) were crossed in half diallel fashion at the experimental farm of Seed and Plant Improvement Institue (SPII), Iran in 2008. The F1 seeds, along with their parents were grown in greenhouse in well water and drought conditions using randomized complete block design with three replications in 2009. Grains of the lines were sown in plastic pots filled with 12 kg of soil composed of a mixture of garden soil, compost and sand (1:1:1, v/v). Well-watered plants were irrigated to water-holding capacity in 3 day intervals.
| Table 1: | List of lines used in the study |
 | |
| DR: Drought resistance, R: Resistant, MR: Moderately resistant, S: Susceptible, GH: Growth habit, W: Winter, F: Facultative, S: Spring | |
The drought stress was started at the mid-flowering stage. Drought was imposed by withholding water until 80% soil moisture depletion and then adding only 20% of the water given to the control pots. Two weeks after onset of drought stress, flag leaves were collected from both control and stress treatments for enzyme assay.
Enzyme extraction and determinations of enzymes activity: Leaf tissues were ground to fine powder in liquid nitrogen, then enzyme extraction was done according to Sairam et al. (1998). Catalase was assayed by the method of Maehly and Chance (1954). The activity of SOD was measured according to the method of Giannopolitis and Ries (1977). The APX activity was assayed by the method of Nakano and Asada (1981). The enzyme activities were expressed in terms of specific activity (Unit/mg Fresh Weight).
Statistical analysis: The diallel analysis was done according to the theoretical basis developed by Hayman (1954a), adapted for the half diallel by Walters and Morton (1978). The goodness of fit of the additive-dominant model was did based on the analysis of variance of Wr-Vr (difference between array parent-offspring covariance and array variance) and linear regression of Wr (array parent-offspring covariance) on Vr (array variance). The genetic components: D, H1, H2, F and h 2 were estimated by method of Singh and Singh (1984). Standard errors of these components were calculated from expected and observed values of Wr, Vr, V r, Vp and (mL1- mL0) over replications. From the estimates of the genetic components, the genetic parameters presented in Table 4 were estimated. Broad sense heritability, narrow sense heritability and average degree of dominance were calculated according to Mather and Jinks (1971). Analysis of variance of diallel was performed using the DIAL98 software Ukai (1989) and genetic components were estimated by electronic spreadsheets in the Excel program (Microsoft® Excel 2003).
RESULTS
In Table 2, it is recognizable that no one from treatments (seven lines) for CAT, APX and SOD was not significant. For CAT, APX and SOD Lowest value was 0.00019 related to Rep (for stress condition in CAT and also, highest value was 1.68 for Rep (Repetition) in stress condition linked to APX (Table 2). The results of the goodness of fit of the additive-dominant model are in Table 2 and 4. The Analysis of variance of the diallel is shown in Table 3. In normal condition all sources of variation in Table 3 for APX and SOD were nonsignificant. In Table 3, highest value was 20.69 for b1 (component which measures the mean deviations of the F1s from the mid-parental values) in stress condition in related to APX and lowest value was 0.00021 for Rep in normal condition about CAT. Non significant Wr-Vr mean squares for treatment indicate the adequacy of additive dominant model for all of traits. The b1 component which measures the mean deviations of the F1s from the mid-parental values was highly significant for SOD and APX activity. For CAT and APX activities in normal condition, the slop of linear regression (b) was significantly lower than unit and additive-dominant model was not satisfied (Table 4).
| Table 2: | Goodness of fit of additive-dominant model based on ANOVA of Wr-Vr |
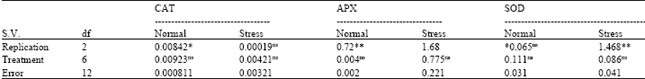 | |
| ns: Non-significant, *and**: Significant at p<0.05 and 0.01, respectively | |
| Table 3: | Analysis of variance of the diallel tables for the evaluated traits |
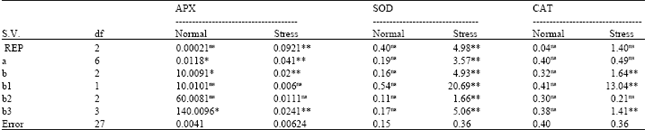 | |
| S.V: Source of variation, ns: Non-significant, *and** significant at p<0.05 and 0.01, respectively. a: Additive variance, b: Dominant genetic effects, b1: Component which measures the mean deviations of the F1s from the mid-parental values, b2: Indicator presence of asymmetry in the distribution of alleles among the parents, b3: Specific combining ability variance | |
| Table 4: | Estimates of genetic components and related statistics in half-diallel design |
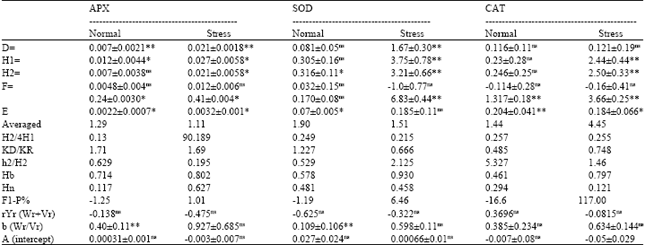 | |
| ns: Bon-significant, *and** significant at p<0.05 and 0.01, respectively, D: Additive genetic variance, H1: Uncorrected dominance genetic variance, H2: Corrected dominancevarianc , F: Average covariation of additive and dominance effects, h: Dominance effects (as algebraic sum over all loci in 2 heterozygous phase in all crosses), Averaged: Average degree of dominance, H2/4 H1: Relative distribution of positive and negative genes among parents, KD/KR: Relative distribution of dominant and recessive genesamong parents, h2/H: The number of effective factors that showed ominance, Hb: Broad senseheritability, Hn: Narrow sense heritability, E: Environmental variance (errormeans quare of simple ANOVA divided by number of replications), rYr( Wr+Vr): Relation between the favorable alleles and dominance, F1-P: Magnitudeofdominance b (Wr/Vr): Slope of regression line of Wr on Vr | |
For CAT and APX activity, additive variance (a component) was highly significant in stress condition, indicating the presence of additive effects in the control of these traits. However, this component was not significant in activity of SOD. The significance of a (additive variance) in Table 2 was elegant with the significance of additive effects (D component) in Table 4. Significance of b1 part was generally in accordance with higher magnitude of dominance ( ) and this indicated that F1s (all hybrids) had higher enzymatic activity than their parents under stress. The b source of variation (dominant genetic effects) also showed highly significant effects for all of traits under stress condition. This proves the importance of dominant genetic effects as well as additive effects in the control of all of traits. The b2 (component that measures average heterosis) section was significant only for APX activity under stress. The proportion of positive and negative genes was calculated by (H2/4H1) in Table 4. This ratio was lower than 0.25 in CAT activity and APX (under stress), indicating the presence of asymmetry in the distribution of the positive and negative alleles in the parents. The b3 component which is synonym with specific combining ability variance was significant for all of traits under stress. Positive values for F substantiated by KD/KR (relative distribution of dominant and recessive genes among parents) being greater than 1 and vice versa. The estimate of the genetic component F was non significant in all cases which is an indication of symmetry in the distribution of dominant and recessive alleles in the parents. However, the ratio of the total number of dominant and recessive alleles in the parents (KD/KR) was higher than 1 for CAT activity, indicates a higher frequency of dominant alleles in the parents, this ratio was lower than 1 in the case of APX (under stress) and SOD activity. For all of traits positively significant h (dominance effects (as algebraic sum over all loci inheterozygous phase in all crosses) alues were recorded (Table 4). The degree of average dominance was higher than 1, indicating the presence of over dominance in control of these chracterss. Contribution of over dominance also confirmed by higher heterosis (117%) in SOD activity (Table 4). The number of groups of genes that control the trait and exhibit dominance (h2/H2) ranged from 0.195 to 5.327 (H2 = corrected dominance variance). In general SOD had the highest number of dominant genes followed by APX and CAT activity. The values of the broad sense heritability (Hb) ranged from 0.461-0.930. The differences observed between the Hn and Hb reflected the presence of the dominant effects. Narrow sense heritability of traits ranged from 0.117 to 0.627. Among the traits CAT activity had the highest Hn, followed by APX with moderate Hn and SOD with low Hn (under stress) and in normal condition APX had highest Hn, followed by SOD with moderate Hn and CAT with low Hn. Non-significant correlation coefficients between the parental means and order of dominance rYr (Wr+Vr) were observed for all characterss indicating that there is not a strong relation between dominance and isotropic of traits.
The degree of average dominance also was shown by the intercept point between origin and regression line. As shown in Table 4, only the intercept of SOD activity (under drought stress) was significantly lower than zero proving its over dominance.
DISCUSSION
Drought stress cause molecular damage to plant cells, that directly or indirectly the reason for the formation of Actived Oxygen Species (AOS). Moreover, it inactivates antioxidant enzymes which are very important for H2O2 scavenging such as catalases (Kono and Fridovich, 1983) and peroxides (Esfandiari et al., 2007). Drought stress increased the superoxide level in cells. If this radical is not scavenged by SOD, it disturbs vital bimolecular (Mittler, 2002). Candan and Tarhan (2003), Martinez et al. (2001), Zhao et al. (2006) and Esfandiari et al. (2007) had similar findings and expressed that the increase in SOD activity and decrease in oxidative damage were closely related. Cloned plant genes and transgenic plants have become a standard tool in plant-stress biology. These technologies have mainly been applied to model systems and have greatly enlarged the knowledge of mechanisms of tolerance. The various abiotic stresses cause changes in plant processes at all levels of organization (morphological, physiological, biochemical and molecular). In recent years, attention has focused on alterations in gene expression. The list of genes whose transcription is upregulated in response to stress is rapidly increasing. Functions for some of these polypeptides are close to being identified and their likely role in stress physiology is being determined. The understanding of mechanisms that regulate gene expression and the ability to transfer genes from other organisms into plants will expand the ways in which plants can be utilized (Joseph and Jini, 2010). Due to the importance of dominance in the control of characters in this experiment, especially APX and CAT activity, it was suggested that the evaluations of lines must be done at advanced generations of inbreeding. Costa et al. (2005) found increase in CAT activity when sorghum plants subjected to 75 mM NaCl and the increase was more conspicuous in tolerant than in sensitive genotype. Sairam et al. (2002) stated that scavenging of H2O2 as represented by GR and CAT is limited and less efficient in susceptible wheat genotypes leading to higher H2O2 accumulation and increasing in lipid peroxidation under water limited environments (Hefny and Abdel-Kader, 2007). Based on the results of this study it was concluded that the activity of antioxidant enzymes under drought stress adequately can be described by additive-dominance model. For APX and CAT activity under stress, additive effects as well as dominant effects were significant. However, in SOD activity, additive effects were not significant. Since the degree of average dominance was higher than 1, the presence of over dominance and greater importance of dominance effects in control of traits was suggested. In general, all of traits had high broad sense heritability and its magnitude was higher under stress than normal conditions but such trend did not observed in narrow sense heritability. Since CAT and APX were controlled by relatively fewer numbers of dominant genes than SOD, it seems that they can be more easily manipulated in plant breeding programs. The results also showed that, effect dominance was predominantly in one direction except for CAT activity, indicating the presence of heterosis in the control of these two traits. These findings get support from previous studies on wheat by Nayyar and Gupta (2006) and Angra et al. (2010). According to Bernardo (2002), individual plant measurements of quantitative traits are prone to large nongenetic effects making estimates of heritability higher. The significance of the b1 component indicates that the dominance was predominantly in one direction and measures average heterosis (Singh and Singh, 1984). The significance of the b2 portion indicated that the mean dominance deviations of the Fs from their mid parental values differed significantly over the F arrays; this proves the presence of asymmetry in the distribution of alleles among the parents (Hayman, 1954b). This means that there was evidence that some parents had a significantly better performance than others (Ramalho et al., 1993). Results obtained about this study from other reviewers are in support ours study but only Saleh and Plieth (2009) reported that the activity of peroxidase has been already reported to be remarkably increased in the case of pea, while its activity is highly decreased in gardencress pepperweed plant (Gholizadeh, 2010).
CONCLUSION
Regarding to the highest heritability of antioxidant enzymes in this study, it can be concluded that they can be considered as good criterion for selecting drought tolerance in tritcale. Since CAT and APX were controlled by relatively fewer numbers of dominant genes than SOD, it seems that they can be more easily manipulated in plant breeding programs. The results also showed that, effect dominance was predominantly in one direction except for CAT activity, indicating the presence of heterosis in the control of these two traits. Antioxidative mechanism seems to be not enough to protect plants from the elevated environmental stresses such as drought. Transgenic plants over expressing single transgene of SOD, APX and GR separately in chloroplast or other compartment of plant cell were generated and displayed increased tolerance against the oxidative stress.
ACKNOWLEDGMENT
The authors would like to thank from SSII for providing the plant material.
REFERENCES
- Ahmad, S., R. Ahmad, M.Y. Ashraf, M. Ashraf and E.A. Waraich, 2009. Sunflower (Helianthus annuus L.) response to drought stress at germination and seedling growth stages. Pak. J. Bot., 41: 647-654.
Direct Link - Amirjani, M.R., 2010. Effect of salinity stress on growth, mineral composition, proline content, antioxidant enzymes of soybean. Am. J. Plant Physiol., 5: 350-360.
CrossRefDirect Link - Angra, S., S. Kaur, K. Singh, D. Pathania, N. Kaur, S. Sharma and H. Nayyar, 2010. Water-deficit stress during seed filling in contrasting soybean genotypes: Association of stress sensitivity with profiles of osmolytes and antioxidants. Int. J. Agric. Res., 5: 328-345.
CrossRefDirect Link - Candan, N. and L. Tarhan, 2003. The correlation between antioxidant enzyme activities and lipid peroxidation levels in Mentha pulegium organs grown in Ca2+, Mg2+, Cu2+, Zn2+ and Mn2+ stress conditions. Plant Sci., 163: 769-779.
CrossRefDirect Link - Chelikani, P., I. Fita and P.C. Loewen, 2004. Diversity of structures and properties among catalases. Cell. Mol. Life Sci., 61: 192-208.
CrossRefDirect Link - Costa, P.H.A., A. Neto, M. Bezerra, J. Prisco and E. Filho, 2005. Antioxidant-enzymatic system of two sorghum genotypes differing in salt tolerance. Braz. J. Plant Physiol., 17: 353-361.
Direct Link - Maehly, A.C. and B. Chance, 1954. The assay of catalases and peroxidases. Methods Biochem. Anal., 1: 357-424.
PubMedDirect Link - Esfandiari, E.O., M.R. Shakiba, S.A. Mahboob, H. Alyari and M. Toorchi, 2007. Water stress, antioxidant enzyme activity and lipid peroxidation in wheat seedling. Int. J. Food Agric. Environ., 5: 149-153.
Direct Link - Giannopolitis, C.N. and S.K. Ries, 1977. Superoxide dismutases. I. Occurrence in higher plants. Plant Physiol., 59: 309-314.
CrossRefPubMedDirect Link - Gholizadeh, A., 2010. Anti-oxidation profile in the leaves of maize inbreds: Elevation in the activity of phenylalanine ammonia lyase under drought stress. J. Plant Sci., 5: 137-145.
CrossRefDirect Link - Habibi, D., M.M.A. Boojar, A. Mahmoudi, M.R. Ardakani and D. Taleghani, 2004. Antioxidative enzymes in sunflower subjected to droughy stress. Proceedings of tha 4th International Crop Science Congress, Sep. 26-Oct. 01, Brisbane, Australia. http://regional.org.au/au/asa/2004/poster/1/3/4/594_habibid.htm.
- Hefny, M. and D.Z. Abdel-Kader, 2007. Antioxidant-enzyme system as selection criteria for salt tolerance in forage sorghum genotypes (Sorghum bicolor L. Moench). Int. J. Plant Breed. Genet., 1: 38-53.
CrossRefDirect Link - Joseph, B. and D. Jini, 2010. Insight into the role of antioxidant enzymes for salt tolerance in plants. Int. J. Bot., 6: 456-464.
CrossRefDirect Link - Kono, Y. and I. Fridovich, 1983. Inhibition and reactivation of Mn-catalase. Implications for valence changes at the active site manganese. J. Biol. Chem., 258: 13646-13668.
PubMedDirect Link - Luna, C.M., G.M. Pastori, S. Driscoll, K. Groten, S. Bernard and C.H. Foyer, 2005. Drought controls on H2O2 accumulation, catalase (CAT) activity and CAT gene expression in wheat. J. Exp. Bot., 56: 417-423.
CrossRefDirect Link - Martinez, C.A., M.E. Loureiro, M.A. Oliva and M. Maestri, 2001. Differential responses of superoxide dismutase in freezing resistant Solanum curtilobum and freezing sensitive Solanum tuberosum subjected to oxidative and water stress. Plant Sci., 160: 505-515.
CrossRefDirect Link - Mittler, R., 2002. Oxidative stress, antioxidants and stress tolerance. Trends Plant Sci., 7: 405-410.
CrossRefPubMedDirect Link - Nakano, Y. and K. Asada, 1981. Hydrogen peroxide is scavenged by ascorbate-specific peroxidase in Spinach chloroplasts. Plant Cell Physiol., 22: 867-880.
CrossRefDirect Link - Pan, Y., L.J. Wu and Z.L. Yu, 2006. Effect of salt and drought stress on antioxidant enzymes activities and SOD isoenzymes of liquorice (Glycyrrhiza uralensis Fisch). Plant Growth Regul., 49: 157-165.
CrossRefDirect Link - Pastori, G.M. and C.H. Foyer, 2002. Common components, networks and pathways of cross-tolerance to stress. The central role of "redox" and abscisic acid-mediated controls. Plant Physiol., 129: 460-468.
PubMedDirect Link - Quiles, M.J. and N.I. Lopez, 2004. Photoinhibition of photosystems I and II induced by exposure to high light intensity during oat plant grown effects on the chloroplastic NADH dehydrogenase complex. Plant Sci., 166: 815-823.
CrossRef - Reddy, A.R., K.V. Chaitanya and M. Vivekanandan, 2004. Drought-induced responses of photosynthesis and antioxidant metabolism in higher plants. J. Plant Physiol., 161: 1189-1202.
CrossRefPubMedDirect Link - Sairam, R.K., P.S. Deshmukh and D.C. Saxena, 1998. Role of antioxidant systems in wheat genotypes tolerance to water stress. Biologia Plantarum, 41: 387-394.
CrossRefDirect Link - Saleh, L. and C. Plieth, 2009. Fingerprinting antioxidative activities in plants. Plant Methods, 5: 2-2.
PubMedDirect Link - Sairam, R.K., K.V. Roa and G.C. Srivastava, 2002. Differential response of wheat genotypes to long term salinity stress in relation to oxidative stress, antioxidant activity and osmolyte concentration. Plant Sci., 163: 1037-1046.
CrossRefDirect Link - Singh, M. and R.K. Singh, 1984. A comparison of different methods of half-diallel analysis. Theor. Applied Genet., 67: 323-326.
CrossRef - Stepien, P. and G. Klobus, 2005. Antioxidant defense in the leaves of C3 and C4 plants under salinity stress. Physiol. Plant., 125: 31-40.
CrossRefDirect Link - Walters, D.E. and J.R. Morton, 1978. On the analysis of variance of a half diallel table. Biometrics, 34: 91-94.
Direct Link - Zhao, F., S. Guo, H. Zhang and Y. Zhao, 2006. Expression of yeast SOD2 in transgenic rice results in increased salt tolerance. Plant Sci., 170: 216-224.
CrossRefDirect Link - Nayyar, H. and D. Gupta, 2006. Differential sensitivity of C3 and C4 plants to water deficit stress: Association with oxidative stress and antioxidants. Environ. Exp. Bot., 58: 106-113.
CrossRefDirect Link