
Wattana Pattanagul
Department of Biology, Faculty of Sciences, Khon Kaen University, Khon Kaen, 40002, Thailand
Asian Journal of Plant Sciences
Year: 2011 | Volume: 10 | Issue: 3 | Page No.: 212-219







ABSTRACT
The objective of this study was to determine the effects of exogenous ABA on growth and carbohydrate metabolism in rice seedlings cv. Khao Dawk Mali 105 under drought stress. Rice seedlings were grown for 10 days and subjected to water restriction for 7 days. Drought stress caused significant reduction in shoot as well as root growth and biomass. Leaf relative water content and chlorophyll contents were reduced by drought stress. Application of exogenous ABA helps the plants by improving leaf relative water content. Drought stress also caused significant increases in sugar accumulation, accompanied by increases in sucrose phosphate synthase activities. Application of exogenous ABA enhanced sugar accumulation but decreased starch content in the leaf. The results indicated that sugar accumulation is associated with drought stress and exogenous abscisic acid can improve plant water status as well as enhances sugar accumulation under drought stress.
PDF Abstract XML References Citation
Received: February 28, 2011;
Accepted: May 22, 2011;
Published: July 28, 2011
How to cite this article
Wattana Pattanagul, 2011. Exogenous Abscisic Acid Enhances Sugar Accumulation in Rice (Oryza sativa L.) under Drought Stress. Asian Journal of Plant Sciences, 10: 212-219.
DOI: 10.3923/ajps.2011.212.219
URL: https://scialert.net/abstract/?doi=ajps.2011.212.219
DOI: 10.3923/ajps.2011.212.219
URL: https://scialert.net/abstract/?doi=ajps.2011.212.219
INTRODUCTION
Water deficit is one of the crucial environmental factors limiting plant growth and productivity and is expected to become increasingly important in many regions because of the ongoing climate change (Xoconostle-Cazares et al., 2010). It is well established that drought stress impairs numerous physiological and biochemical processes in plants. Photosynthetic rate is heavily reduced under drought condition due to stomatal closure, so it is assumed to be responsible for decreased dry matter production (Sepehri and Modarres Sanavy, 2003; Lawlor and Tezara, 2009). The production of Reactive Oxygen Species (ROS) can seriously disrupt normal metabolic process during stress through chlorophyll loss, membrane lipid peroxidation, protein carbonylation, inactivating the -SH containing enzymes and ultimately cell death (Farooq et al., 2009b; Basu et al., 2010). Therefore, by limiting plant growth, production and consumption of photoassimilates will inevitably be altered at both the leaf and the whole plants levels (Praxedes et al., 2006). Plants can response and tolerate water stress by altering their cellular metabolism and evoking various defense mechanisms including stomatal closure, accumulation of solutes, cell wall hardening and production of proteins and enzymes involved in cellular protection and ROS scavenging (Guo et al., 2006; Esfandiari et al., 2008; Omidi, 2010).
Abscisic Acid (ABA) has been shown to mediate many physiological and developmental processes throughout the life cycle of plants including responses of plants to environmental stresses. Its level increase as a result of stresses including drought, salinity and cold stresses that involved cellular water stress (Khadri et al., 2006). ABA is known to act as a major signaling molecule involved in the response of plants to drought stress. Stress-related responses induced by ABA often occur earlier than the change of plant water status during soil drying and thereby constitute the first line of defense as soil water deficits are encountered (Liu et al., 2005). The hormone triggers stomatal closure to limit water loss through transpiration, as well as mobilizes a battery of genes that presumably serve to protect the cells from the ensuing oxidative damage in prolonged stress (Wasilewska et al., 2008). These physiological and biochemical changes have been proposed to reduce the deleterious effects induced by water stress. Many studies have shown that ABA is able to induce changes including synthesis of stress proteins, proline, sugar alcohols, soluble carbohydrates and glycine betaine in which may involve in stress tolerance (Bagniewska-Zadworna et al., 2007).
Carbohydrate metabolism is strongly affected by drought stress. Water stressed plants often accumulate sugars and their derivatives, such as polyols and raffinose family oligosaccharides (Valliyodan and Nguyen, 2006; Toldi et al., 2009). Accumulation of these osmolytes may help plants to tolerate dehydration by improving their ability to maintain osmotic balance within the cell (Choluj et al., 2008; Costa et al., 2008). Additional benefits of these solutes have been described, including buffering cellular redox potential, protecting the cell from dehydration by stabilizing membrane and protein structures and providing possible energy source under severe stress (Hasegawa et al., 2000). Furthermore, alteration in photoassimilate partitioning between source and sink tissues may also contribute to the accumulation of these solutes (Hare et al., 1998). The aim of this study was to investigate the effects of exogenous ABA on growth and carbohydrate metabolism in rice seedling under drought stress. The results could be important for better understanding the mechanism of drought tolerance in rice.
MATERIALS AND METHODS
Plant material, growth conditions and treatment: Rice (Oryza sativa L.) seeds, cvs. Khao Dawk Mali 105 was obtained from Rice Research Station, Khon Kaen, Thailand. Seeds were soaked in 5% sodium hypochlorite for 15 min and rinsed thrice with distilled water. Rice seedlings were germinated by placing the seeds on the moistened filter paper for 3 days. Approximately 10 seedlings were then potted in a plastic pot containing soil:sand:peat mix (1:1:2, v/v) and kept in the greenhouse at the Department of Biology, Faculty of Sciences, Khon Kaen University, Thailand under natural illumination and temperature conditions. The seedlings were watered daily with 100 mL half strength Hoagland solution and allowed to grow for 10 days. The seedlings were then separated into 4 treatments containing 5 pots per treatment. The control plants were sprayed to runoff with distilled water and watered daily with 100 mL distilled water. For the drought stress treatment, the plants were sprayed to runoff with distilled water, one day prior to drought treatment and water was withheld for a period of 7 days. For exogenous ABA treatment, the seedlings were sprayed to runoff with an aqueous solution of ABA (20 mg L-1) and watered daily with 100 mL distilled water. For exogenous ABA and drought treatment, the seedlings were sprayed to runoff with an aqueous solution of ABA (20 mg L-1), one day prior to drought treatment and water was withheld for a period of 7 days thereafter.
Plant growth, chlorophyll and relative water content measurements: After drought treatment, the seedlings were randomly sampled from each pot. Root and shoot lengths of 5 plants from each treatment were determined. Fresh weights of shoot and root tissues were recorded and then the tissues were oven-dried at 80°C for 3 days in order to determine dry weight. Five additional plants from each treatment were randomly selected to determine chlorophyll and leaf relative water contents. About 0.1 g leaf sample was used for chlorophyll determination. Leaf samples were extracted with 5 mL of acetone and chlorophyll contents were determined spectrophotometrically at 645 and 663 nm (Arnon, 1949). For leaf relative water content determination, about 0.1 g leaf sample was cut into smaller pieces and weighed to determine fresh weight. The leaf sample was floated in freshly de-ionized water for 12 h and weighed thereafter to determine fully turgid weight. The leaf sample was oven-dried at 80°C for 3 days and the dry weight was obtained. The Relative Water Content (RWC) was determined using the following formula:
where, FW is fresh weight, DW is dry weight and tW is Turgid weight (Turner, 1981).
Carbohydrate extraction and analysis: Fully mature leaves were randomly selected from each treatment and frozen on dry ice to transfer to the laboratory. Approximately 0.1 g leaf tissue was used for carbohydrate analysis. Soluble sugars were extracted from the tissues in hot 80% (v/v) ethanol and determined as previously described (Pattanagul and Thitisaksakul, 2008). Starch in the leaf residue was digested with amyloglucosidase overnight and released glucose was quantified enzymically using hexokinase/glucose-6-P dehydrogenase.
Enzyme extraction and assay: About 1 g leaf tissue was collected during midday for enzyme extraction and determination of sucrose phosphate synthase and invertase activities. The leaf sample was ground on ice using mortar and pestle in 5 mL grinding buffer containing 50 mM HEPES, 10 mM MgCl2, 1 mM EDTA, 0.25% BSA and 5 mM dithiothreitol, pH 7.5. The extract was filtered through 4 layers of cheesecloth, then centrifuge for 1 min at 10000 g. Crude extract was desalted on 2 mL Sephadex G25 columns equilibrated with the grinding buffer. The amount of protein in the enzyme extract was determined by Bradford method (Bradford, 1976).
SPS activity was determined by measuring sucrose-6-phosphate produced from the substrates, UDP-glucose and fructose-6-phosphate. Approximately 80 μL desalted enzyme was incubated in a reaction mixture containing 25 mM UDP-glucose, 8 mM fructose-6-phosphate, 5 mM MgCl2 at 25°C for 1 h and terminated by adding 100 μL of 1 N NaOH. Unreacted fructose-6-phosphate and fructose were destroyed by boiling the tube in a boiling water bath for 10 min. Sucrose-6-phosphate formed during the reaction was determined by reacting with 0.25 mL resorcinol solution and quantified by a spectrophotometer at 520 mM (Robbins and Pharr, 1987).
Invertase activities were determined under acidic (pH 4.0) and alkaline (pH 7.6) conditions. Approximately 50 μL of desalted enzyme was incubated with 125 mM sucrose (w/v) in the extraction buffer at the appropriate pH. Assays were run at 30°C for 30 min and stopped by boiling for 1 min. The glucose content of 10 μL aliquots of the assay mixture was determined spectrophotometrically at 340 nm using hexokinase/glucose-6-P dehydrogenase enzymes.
Data analysis: Statistical analyses were carried out by ANOVA tests with the SPSS program version 17.0 (SPSS Inc., Chicago, IL). Significant differences were determined by post-hoc comparison (Duncan’s multiple range test) at p<0.05.
RESULTS
Plant growth, chlorophyll and relative water contents: Drought stress caused significant reduction in shoot growth. Shoot length was reduced from 40.20 cm plant-1 in the control plants to 31.40 cm plant-1 in the droughted plants (Fig. 1a). Shoot fresh and dry weights were also reduced from 0.34 and 57.28 mg plant-1 in the control plants to 0.11 and 35.2 mg plant-1 in the drought plants (Fig. 1c, e). Shoot fresh weights were heavily reduced by drought stress accompanied by the significance reduction in leaf relative water content. Exogenous ABA also caused reduction in shoot growth, similar to the effects of drought stress but to the lesser extends (Fig. 1c). In addition, application of exogenous ABA also showed similar effects in the shoot tissue as seen in the droughted plants. On the other hand, root lengths were not significantly affected by drought stress. Both root fresh and dry weights, however, were significantly reduced under drought stress. Root fresh and dry weights were reduced from 0.09 and 11.52 mg plant-1 in the control plants to 0.02 and 6.46 mg plant-1 in the drought plants (Fig. 1d, f).
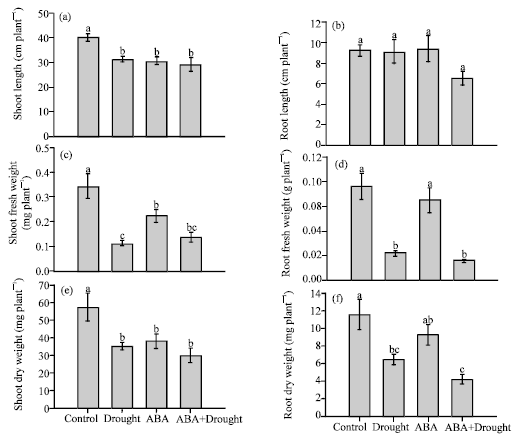 | |
| Fig. 1(a-f): | Effects of drought and exogenous ABA on shoot length (a), root length, (b) shoot fresh weight, (c) root fresh weight and (d) shoot dry weight (e) and root dry weight (f) in mature leaves. Data represent the Means±SE of five measurements per data point. Values showing the same letter are not significantly different |
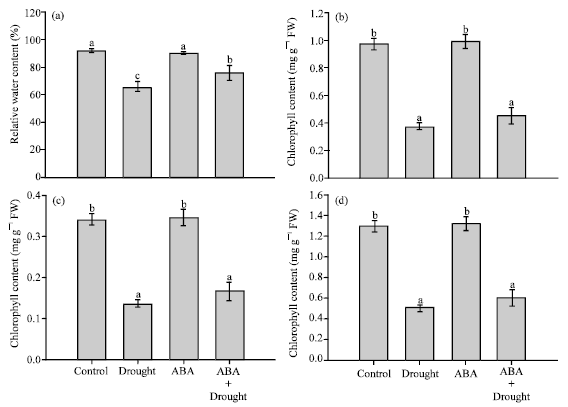 | |
| Fig. 2 (a-d): | Effects of drought and exogenous ABA on (a) relative water content, (b) chlorophyll a, (c) chlorophyll b, (d) and total chlorophyll in mature leaves. Data represent the Means±SE of five measurements per data point. Values showing the same letter are not significantly different |
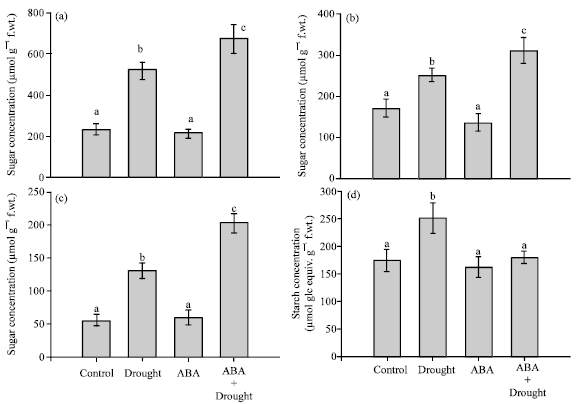 | |
| Fig. 3 (a-d): | Effects of drought and exogenous ABA on (a) total soluble sugar, (b) sucrose, (c) fructose, (d) and starch (d) in mature leaves. Data represent the Means±SE of five measurements per data point. Values showing the same letter are not significantly different |
In contrast to the shoot tissue, application of exogenous ABA to the unstressed plant caused no reduction in the root biomass.
Leaf relative water content was significantly reduced by drought stress. Leaf relative water content was reduced from 91.89% in the control plants to 65.73% in the drought plant (Fig. 2a). Exogenous ABA had no effects in leaf relative water content in the unstressed plants. On the other hand, exogenous ABA increased relative water content in the droughted plants to 75.93% compared to 65.73% in the droughted plants (Fig. 2a). In addition, drought stress also caused reduction in chlorophyll a, b and total chlorophyll contents. Chlorophyll a, b and total chlorophyll were reduced from 0.97, 0.34 and 1.31 mg g-1 FW, respectively, in the control plants to 0.37, 0.13 and 0.51 mg g-1 FW, respectively, in the droughted plants (Fig. 2b, d). Although application of exogenous ABA showed slight increases in chlorophyll contents compared to the droughted plants, the increases were not significantly different.
Carbohydrate contents: Water deficit stress caused a significant increase in leaf total soluble sugars. Total soluble sugar was increased from 232.40 μmol g-1 FW in the control plants to 623.98 μmol g-1 FW in the droughted plants (Fig. 3a). Sucrose and fructose were also increased similarly (Fig. 3b, c). Application of exogenous ABA also resulted in a higher increase in total soluble sugars compared to the droughted plants. Exogenous ABA caused increase in leaf total soluble sugar, sucrose and fructose to 672.89, 309.65 and 203.18 μmol g-1 FW, respectively. Application of exogenous ABA to the unstressed plants, however, showed no difference in total soluble sugar compared to the control plants. Water restriction also resulted in an increase in leaf starch content (Fig. 3d). On the contrary, exogenous ABA showed no increase in starch accumulation compared to the control plants (Fig. 3d).
Enzyme activities: Accumulation of sugars in the stressed plants was also reflected in an increase in SPS activities. SPS activities increased from 0.66 μmol product mg-1 protein h-1 in the control plants to 1.11 μmol product mg-1 protein h-1 in the droughted plants (Fig. 4a). Exogenous ABA also caused increases in SPS activities to 0.96 μmol product mg-1 protein h-1. In addition, application of exogenous ABA to the unstressed plants had no effect on SPS activities.
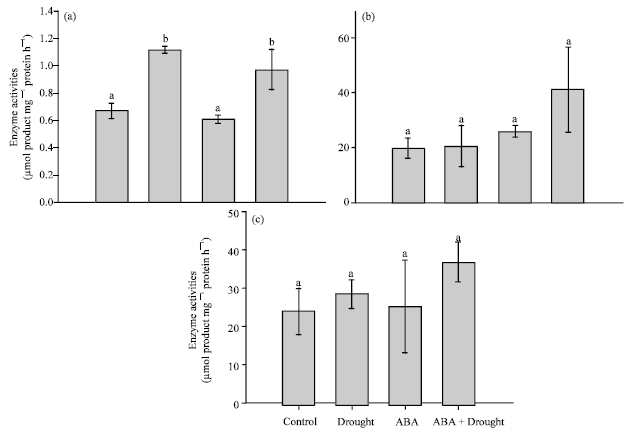 | |
| Fig. 4 (a-c): | Effects of drought and exogenous ABA on activities of (a) sucrose phosphate synthase, (b) acid invertase, (c) and alkaline invertase, (d) in mature leaves. Data represent the Means±SE of five measurements per data point. Values showing the same letter are not significantly different |
On the other hand, water deficit stress and ABA had no significant effect on both acidic and alkaline invertase activities (Fig. 4b, c).
DISCUSSION
Upon water deficit stress, the seedling growth was largely decreased. This may be partly due to lower turgor pressure and decreased photosynthetic rate in the cells (Cha-Um et al., 2007; Regier et al., 2009). Exogenous ABA also caused reduction in shoot growth, similar to drought condition. The hormone has been known to trigger stomatal closure in order to limit water loss under water stress, thus limiting photosynthetic CO2 assimilate (Liu et al., 2005). Exogenous ABA was also reported to reduce photosynthetic rate, stomatal conductance and transpiration rate in cotton (Pandey et al., 2003).
The reduction in leaf relative water content was provoked by the water deficiency in soil (Hassanzadeh et al., 2009). Exogenous ABA helps the plants to better maintain cellular water level which were also reported in chickpea, Ilex paraguariensis and Polypodium vulgarei (Sansberro et al., 2004; Bagniewska-Zadworna et al., 2007; Kumar et al., 2008). In this study, application of exogenous ABA resulted in increased sugar accumulation which may partly responsible for improving leaf relative water content. In addition, application of ABA are known to affect plant growth and development, mimicking the effects of water stress, thereby helping plants to better survive stress conditions (Farooq et al., 2009a).
Under drought condition, sugar accumulation was observed. The increase of sucrose was accompanied by significance increase in SPS activities. Stressed-induced overproduction of sugars has been reported in numerous species and is believed to play an important role in drought tolerance. In soybean, total soluble carbohydrates, sucrose and reducing carbohydrates were increased under water deficit (Adejare and Umebese, 2008; Lobato et al., 2008). In robusta coffee (Coffea canephora Pierre var. kouillou) leaves, drought tolerance clones showed increases in hexoses and sucrose in response to drought whereas their levels in drought sensitive clones remained unchanged (Praxedes et al., 2006). Accumulation of sugars can help to maintain osmotic balance as well as protect enzymes and membrane against deleterious effects of destabilizing ions (Farooq et al., 2009a). Furthermore, application of exogenous ABA even triggers more sugar accumulation, but decreases starch content. ABA also causes increases in sugar accumulation as well as decreased starch content in Polypodium vulgare (Bagniewska-Zadworna et al., 2007). It has been suggested that under drought stress, the products from starch hydrolysis could be the substrate for sucrose synthesis (Lee et al., 2008). ABA-induced sugar accumulation may partly increase osmotic adjustment in which helps the plants to better survive under drought condition.
In conclusion, this study demonstrates that accumulation of sugars was associated with drought stress. Accumulation of sugar is considered to play an important role in drought tolerance. Exogenous ABA was also shown to help the plants to maintain their relative water content and enhanced sugar accumulation under drought stress.
ACKNOWLEDGMENTS
This research is supported in part by Khon Kaen University Research Fund (No. 523701).
REFERENCES
- Adejare, F.B. and C.E. Umebese, 2008. Water stress induces cultivar dependent changes in stomatal complex, yield and osmotic adjustments in Glycine max L. Int. J. Agric. Res., 3: 287-295.
CrossRefDirect Link - Arnon, D.I., 1949. Copper enzymes in isolated chloroplasts. Polyphenoloxidase in Beta vulgaris. Plant Physiol., 24: 1-15.
CrossRefPubMedDirect Link - Bagniewska-Zadworna, A., E. Zenkteler, K. Czaczyk and M. Osinska, 2007. The effect of dehydration with or without abscisic acid pretreatment on buds regeneration from Polypodium vulgare L. rhizomes. Acta Physiol. Plant., 29: 47-56.
CrossRef - Basu, S., A. Roychoudhury, P.P. Saha and D.N. Sengupta, 2010. Differential antioxidative responses of indica rice cultivars to drought stress. Plant Growth Regul., 60: 51-59.
CrossRefDirect Link - Bradford, M.M., 1976. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem., 72: 248-254.
CrossRefPubMedDirect Link - Cha-Um, S., S. Roytakul, T. Sathung, A. Maijandang and C. Kirdmanee, 2007. Effect of exogenous glucose and abscisic acid on physiological and morphological performances of in vitro indica rice (Oryza sativa L. sp. indica). Am. J. Plant Physiol., 2: 155-166.
CrossRefDirect Link - Choluj, D., R. Karwowska, A. Ciszewska and M. Jasińska, 2008. Influence of long-term drought stress on osmolyte accumulation in sugar beet (Beta vulgaris L.) plants. Acta Physiol. Plant., 30: 679-687.
Direct Link - Costa, R.C.L., A.K.S. Lobato, C.F.O. Neto, P.S.P. Maia, G.A.R. Alves and H.D. Laughinghouse, 2008. Biochemical and physiological responses in two Vigna uguiculata (L.) walp. Cultivars under water stress. J. Agron., 7: 98-101.
CrossRefDirect Link - Esfandiari, E., M.R. Shakiba, S.A. Mahboob, H. Alyari and S. Shahabivand, 2008. The effect of water stress on the antioxidant content, protective enzyme activities, proline content and lipid peroxidation in wheat seedling. Pak. J. Biol. Sci., 11: 1916-1922.
CrossRefPubMedDirect Link - Farooq, M., S.M.A. Basra, A. Wahid, N. Ahmad and B.A. Saleem, 2009. Improving the drought tolerance in rice (Oryza sativa L.) by exogenous application of salicylic acid. J. Agron. Crop Sci., 195: 237-246.
CrossRefDirect Link - Guo, Z., W. Ou, S. Lu and Q. Zhong, 2006. Differential responses of antioxidative system to chilling and drought in four rice cultivars differing in sensitivity. Plant Physiol. Bioch., 44: 828-836.
CrossRef - Hare, P.D., W.A. Cress and J. van Staden, 1998. Dissecting the roles of osmolyte accumulation during stress. Plant Cell Environ., 21: 535-553.
CrossRefDirect Link - Hasegawa, P.M., R.A. Bressan, J.K. Zhu and H.J. Bohnert, 2000. Plant cellular and molecular responses to high salinity. Annu. Rev. Plant Physiol. Plant Mol. Biol., 51: 463-499.
CrossRefDirect Link - Hassanzadeh, M., A. Ebadi, M. Panahyan-e-Kivi, A.G. Eshghi, Sh. Jamaati-e-Somarin, M. Saeidi and R. Zabihi-e-Mahmoodabad, 2009. Evaluation of drought stress on relative water content and chlorophyll content of sesame (Sesamum indicum L.) genotypes at early flowering stage. Res. J. Environ. Sci., 3: 345-350.
CrossRefDirect Link - Khadri, M., N.A. Tejera and C. Llunch, 2006. Alleviation of salt stress in common bean (Phaseolus vulgaris) by exogenous abscisic acid supply. J. Plant Growth Regul., 25: 110-119.
Direct Link - Kumar, S., G. Kaur and H. Nayyar, 2008. Exogenous application of abscisic acid improves cold tolerance in chickpea (Cicer arietinum L.). J. Agron. Crop Sci., 194: 449-456.
CrossRef - Lawlor, D.W. and W. Tezara, 2009. Causes of decreased photosynthetic rate and metabolic capacity in water-deficient leaf cells: A critical evaluation of mechanisms and integration of processes. Ann. Bot., 103: 561-579.
CrossRefPubMedDirect Link - Liu, F., C.R. Jensen and M.N. Andersen, 2005. A review of drought adaptation in crop plants: changes in vegetative and reproductive physiology induced by ABA-based chemical signals. Aust. J. Agr. Res., 56: 1245-1252.
Direct Link - Lobato, A.K.S., C.F.O. Neto, R.C.L. Costa, B.G.S. Filho, F.J.R. Cruz and H.D. Laughinghouse, 2008. Biochemical and physiological behavior of Vigna unguiculata (L.) Walp. Under water stress during the vegetative phase. Asian J. Plant Sci., 7: 44-49.
CrossRefDirect Link - Omidi, H., 2010. Changes of proline content and activity of antioxidative enzymes in two canola genotype under drought stress. Am. J. Plant Physiol., 5: 338-349.
CrossRefDirect Link - Pandey, D.M., C.L. Goswami and B. Kumar, 2003. Physiological effects of plant hormones in cotton under drought. Biol. Plant., 47: 535-540.
Direct Link - Pattanagul, W. and M. Thitisaksakul, 2008. Effects of salinity stress on growth and carbohydrate metabolism in three rice (Oryza sativa L.) cultivars differing in salinity tolerance. Indian J. Exp. Biol., 46: 736-742.
PubMed - Praxedes, S.C., F.M. DaMatta, M.E. Loureiro, M.A.G. Ferrao and A.T. Cordeiro, 2006. Effects of long-term soil drought on photosynthesis and carbohydrate metabolism in mature robusta coffee (Coffea canephora Pierre var. kouillou) leaves. Environ. Exp. Bot., 56: 263-273.
Direct Link - Regier, N., S. Streb, C. Cocozza, M. Schaub, P. Cherubini, S.C. Zeeman and B. Frey, 2009. Drought tolerance of two black poplar (Populus nigraL.) clones: Contribution of carbohydrates and oxidative stress defence. Plant Cell Environ., 32: 1724-1736.
Direct Link - Robbins, N.S. and D.M. Pharr, 1987. Regulation of photosynthetic carbon metabolism in cucumber by light intensity and photosynthetic period. Plant Physiol., 85: 592-597.
CrossRefDirect Link - Sansberro, P.A., L.A. Mroginski and R. Bottini, 2004. Foliar sprays with ABA promote growth of Ilex paraguariensis by alleviating diurnal water stress. Plant Growth Regul., 42: 105-111.
Direct Link - Sepehri, A. and S.A.M.M. Sanavy, 2003. Water and nitrogen stress on maize photosynthesis. J. Biol. Sci., 3: 578-584.
CrossRefDirect Link - Toldi, O., Z. Tuba and P. Scott, 2009. Vegetative desiccation tolerarnce: Is it a goldmine for bioengineering crops. Plant Sci., 176: 187-199.
Direct Link - Turner, N.C., 1981. Techniques and experimental approaches for the measurement of plant water status. Plant Soil, 58: 339-366.
CrossRefDirect Link - Valliyodan, B. and H.T. Nguyen, 2006. Understanding regulatory networks and engineering for enhanced drought tolerance in plants. Curr. Opin. Plant Biol., 9: 189-195.
PubMed - Wasilewska, A., F. Vlad, C. Sirichandra, Y. Redko and F. Jammes et al., 2008. An update on abscisic acid signaling in plants and more. Mol. Plant, 1: 198-217.
CrossRefPubMedDirect Link - Xoconostle-Cazares, B., F.A. Ramirez-Ortega, L. Flores-Elenes and R. Ruiz-Medrano, 2010. Drought tolerance in crop plants Am. J. Plant Physiol., 5: 241-256.
CrossRef - Lee, B., Y. Jin, W. Jung, J. Avice, A. Morvan-Bertrand, A. Ourry, C. Park and T. Kim, 2008. Water-deficit accumulates sugars by starch degradation-not by de novo synthesis-in white clover leaves (Trifolium repens). Physiol. Plant., 134: 403-411.
CrossRef - Farooq, M., A. Wahid, O. Ito, D.J. Lee and K.H.M. Siddique, 2009. Advances in drought resistance of rice. Crit. Rev. Plant Sci., 28: 199-217.
CrossRef