
Shiou Yih Lee
Forest Biotech Laboratory, Department of Forest Management, Faculty of Forestry, Universiti Putra Malaysia, 43400 UPM Serdang, Selangor, Malaysia
Jean Weber
Bio-Resource Management Lab, Institute of Tropical Forestry and Forest Products (INTROP), Universiti Putra Malaysia, 43400 UPM Serdang, Selangor, Malaysia
Rozi Mohamed
Forest Biotech Laboratory, Department of Forest Management, Faculty of Forestry, Universiti Putra Malaysia, 43400 UPM Serdang, Selangor, Malaysia
Asian Journal of Plant Sciences
Year: 2011 | Volume: 10 | Issue: 3 | Page No.: 202-211







ABSTRACT
Aquilaria (Thymelaeaceae) is an endangered agarwood-producing tropical tree that is endemic to the Indomalesia region. Molecular information on genetic diversity of Aquilaria is limited. The aims of this research were to study genetic diversity among three Aquilaria species, growing in natural and distant populations in Malaysia using RAPD markers and to develop SCAR markers for easy identification of A. malaccensis, the major agarwood producer. By analyzing 23 RAPD primers, a total of 368 bands were scored. Multi-populations Descriptive statistics revealed that 333 (90.49%) polymorphic bands were found at species level, where A. malaccensis had 107 (29.08%) bands, A. hirta had 56 (15.22%) and Aquilaria sp.1 had 11 (2.99%), for the percentage of polymorphic loci in a single population/species. Nei’s unbiased measurement indicated moderate similarities among populations/species. Out of the 23 RAPD primers, three were found specific to A. hirta and one was specific to each A. malaccensis and Aquilaria sp.1. RAPD-based SCAR markers generated a total of five species-specific amplicons: three for A. hirta, one each for A. malaccensis and Aquilaria sp.1. SCAR markers for A. malaccensis were used to distinguish five other different A. malaccensis populations in Malaysia. SCAR markers for A. malaccensis tested in five other different A. malaccensis populations in Malaysia yielded positive and consistent results. The DNA fingerprints identified for each Aquilaria sp. will be useful for Aquilaria identification in natural population, young plantation and even at seedling and seed stages in the nursery, as it is rapid and cost-effective and does not rely on morphology.
PDF Abstract XML References Citation
Received: April 04, 2011;
Accepted: May 30, 2011;
Published: July 28, 2011
How to cite this article
Shiou Yih Lee, Jean Weber and Rozi Mohamed, 2011. Genetic Variation and Molecular Authentication of Selected Aquilaria Species from Natural Populations in Malaysia Using RAPD and SCAR Markers. Asian Journal of Plant Sciences, 10: 202-211.
DOI: 10.3923/ajps.2011.202.211
URL: https://scialert.net/abstract/?doi=ajps.2011.202.211
DOI: 10.3923/ajps.2011.202.211
URL: https://scialert.net/abstract/?doi=ajps.2011.202.211
INTRODUCTION
Aquilaria, a genus from the family of Thymelaeaceae, is an endangered woody tree in Malaysia. It is endemic to the Indomalesia region (Mabberley, 2008). The tree is often harvested for its highly valuable fragrant wood called “gaharu” or “agarwood” which the tree produces upon pathological infection or mechanical wounding. Agarwood is highly prized in both local and foreign markets because of its usage in traditional medicines and other products (Barden et al., 2000; Naef, 2011). Five different Aquilaria species have been recorded in Malaysia, including A. hirta Ridl., A. beccariana Van. Tiegh., A. rostrata Ridl., A. malaccensis Lam. and A. microcarpa Baill. which are distributed in the Peninsular and East Malaysia (Chua, 2008). This endemic genus with restricted distributions has received concerns from foresters, biologists and naturalists who aim at conserving these species in the wild. Due to the uncertainty regarding the tree size or age that contain agarwood, harvesting process often involve indiscriminate felling of both infected and uninfected trees in the wild which in turn threaten the habitat of these trees (CITES, 2004). Overexploitation of the species in their natural environments is expected to be reduced by the development of massive ex-situ plantations combined with techniques able to induce agarwood production on young plants (Faridah-Hanum et al., 2009). Yet, the production and breeding of Aquilaria in the nursery require tools for the identification of seeds and seedlings. For Aquilaria in general, species recognition relies mainly on the leave, ovary, fruit and calyx characteristics (Tawan, 2004) and remains subjective if seedling distributions cannot be related to the position of an adult tree in the wild. At the nursery, possibilities for morphological identification are further degraded with changes in the expression of morphologies such as the leave size that is affected by variation of environmental conditions (personal observation). Sequence-based species identification using the plastid trnL-trnF intergenic spacer and nuclear ribosomal ITS regions have been reported for Aquilaria (Eurlings and Gravendeel, 2005; Kiet et al., 2005). However, in the context of production, resorting to DNA sequences or even Restriction Fragment Length Polymorphism (RFLP) (Jena et al., 2009; Ridgway et al., 2003) is much costlier than Random Amplified Polymorphic DNA (RAPD) and microsatellite DNA which in most cases, needs only standard PCR followed by simple agarose electrophoresis. Besides financial and technical constrains the choice of identification methods depends on the genetic resolution needed. Microsatellite markers or Simple Sequence Repeat (SSR) markers have for example been developed for populations studies of A. sinensis in China (Zhang et al., 2009) and A. crassna in Vietnam (Eurlings et al., 2010). The aptitude of SSR markers to reveal hyper variable DNA length polymorphism makes them ideal for detecting population’s genotypic variability and for revealing segregations of alleles in breeding programs or wild population dynamics studies (Ouborg et al., 1999; Patra et al., 2011). However, in comparison to the design of RAPD primers, the high labor required for the development of SSR primers reduces the opportunities of finding adequate markers for cross-species amplifications. RAPD markers have a large coverage of the genome and can rapidly produce high number of polymorphisms. They have for example been used for the identification of grapevine, tomato and peach (Gokbayrak and Soylemezoglu, 2010; Kulkarni and Deshpande, 2006; Melgoza-Villagomez et al., 2009) and are consistent for cultivar registrations (Al-Saghir and Abdel Salam, 2011; Linos et al., 2002). Conversely, in some cases, fidelity of the RAPD as a genetic marker has been questioned since its reproducibility can be altered by small variation of DNA quality or primer and template concentration (Theerakulpisut et al., 2008). In order to overcome these limitations, SCAR (Sequence Characterized Amplified Regions) markers have been developed to contain longer primers from the sequence of a cloned RAPD fragment (Paran and Michelmore, 1993). In this study, RAPD technique was used to detect molecular polymorphisms and genetic variations among selected Aquilaria species in Malaysia. At the same time specific SCAR primers were designed and tested which allowed for a simple and rapid identification of these species at the nursery.
MATERIALS AND METHODS
Plant materials: For genetic diversity analysis and marker development, tree saplings were collected from three natural populations and grown in poly bags in the nursery under natural environment at the Faculty of Forestry, Universiti Putra Malaysia, Selangor, Malaysia. Young leaves from these saplings were collected for genomic DNA isolation. Sampling was conducted in 2009 to 2010 when young shoots became abundant. The three Aquilaria species were: 1) A. hirta from the Merchang Reserve Forest provided by the Forestry Department of Terengganu, 2) A. malaccensis from Karak, provided by the National Seed and Planting Material Procurement Centre at Lentang, Pahang and 3) Aquilaria sp.1 from the Botanical Research Centre of Sarawak Forestry Corporation, Semenggoh, Sarawak. Both A. malaccensis and A. hirta were represented by six individuals while Aquilaria sp.1 was represented by only two individuals because of its scarcity. There is a possibility that the individuals were from a half-sib family as they were collected from the vicinity of a single mother. For testing the specificity and consistency of the SCAR markers, individual A. malaccensis trees were collected from five natural populations in Peninsular Malaysia including from Bukit Bauk (Terengganu), Gua Musang (Kelantan), Jelebu (Negeri Sembilan), Jeli (Kelantan) and Sungai Udang (Melaka). Geographical location of each population is shown in Fig. 1.
RAPD assay: Plant genomic DNA was extracted from 2 g of young leaves and shoots using the DNeasy® Plant Mini Kit (Qiagen, Germany), according to the manufacturer’s protocol, with minor modifications. The quantity and quality were determined by spectrophotometry (NanoPhotometer™, IMPLEN, Germany). For PCR amplifications, genomic DNA samples were diluted to 50 ng μL-1. A total of 60 10-base RAPD primers (Eurofins MWG Operon, USA) from series A, B and C were initially screened for amplification. Twenty-three primers which generated clear and reproducible banding patterns were chosen for the final RAPD analysis. PCR amplification was carried out according to the method described by Williams et al. (1990), with minor modifications. The final reaction volume was 15 μL and contained: 0.67 x PCR buffer, 0.08 mM MgCl2, 0.02 mM of each dNTPs, 1 U of Taq DNA polymerase (Invitrogen, USA), 0.5 mM of primer (Eurofins MWG Operon, USA) and 50 ng of genomic DNA template. Negative controls with distilled water instead of DNA were included in each run in order to verify the absence of contamination. DNA amplification was conducted in a MyCycler™ Thermalcycler (Bio-Rad, USA), programmed for 3 min at 94°C; 35 cycles for 54 sec at 94°C, 43 sec at 45°C and 2 min at 72°C, with a final 5 min extension at 72°C. Amplification products were separated using electrophoresis on 2% agarose gels in 1 X TAE buffer, stained in ethidium bromide (0.5 μg mL-1) and photographed under UV light (FlourChem™ Bio-imaging System, Alpha Innotech, USA).
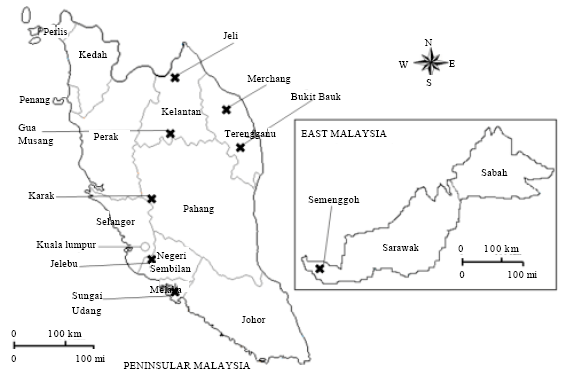 | |
| Fig. 1: | Locations of Aquilaria populations in Malaysia, from which samples were collected. Crosses identify the populations which were assessed in the study |
A 1 kbp DNA ladder (Fermentes, Canada) was included in the gels as a size reference. PCR amplification was repeated at least twice to show reproducibility of the results.
Statistical analysis: The RAPD profiles were scored visually and with the aid of the gel documentation system. Only clear and distinct RAPD bands were scored. Two binary matrices based on the RAPD banding patterns were created, where “1” was attributed to the presence of the targeted loci and “0” to the absence of a band. A similarity index was used to compare the banding patterns between the population/species. Nei’s unbiased genetic distances (Nei, 1978) were calculated among populations using the POPGENE software (version 1.32) (Yeh et al., 1997). A dendrogram was constructed to show the genetic distance between different species in this study using the Unweighted Paired Group of Arithmetic Means Analysis (UPGMA) using the NTSYS-pc programme version 2.01e (Rohlf, 1990).
Cloning and sequencing of RAPD fragments: PCR was repeated three times for each DNA sample. Only reproducible bands below 1 kbp were considered for cloning. The species-specific amplicons were poked with pipette tips and dipped into freshly-prepared PCR mixture with the final reaction volume of 15 μL. All other components were as described above. The PCR mixture was re-amplified until a clear single band was observed. The specific band was excised from the ethidium bromide stained gel under UV light and purified using QIAquick Gel Extration Kit (QIAGEN®, Germany). The recovered DNA fragment was cloned into the pGEM® T-Easy vector (Promega®, USA). The ligation reaction contained 5 μL of 2X Rapid ligation buffer, 1 μL pGEM® T-Easy vector (50 ng), 3 μL PCR product and 1 μL T4 DNA Ligase (3 Weiss units μL-1) in a total volume of 10 μL. The reaction was incubated overnight at 4°C. The ligation reaction was then transformed into Escherichia coli strain JM109 competent cells, by adding 25 μL recently thawed competent cells to a sterile 1.5 mL microcentrifuge tube containing 2 μL of the ligation reaction on ice. Recombinants were identified as white colonies on LB plates with X-gal and IPTG. Positive transformants were screened for gene insert using PCR method. Upon selection, the white colony was grown in 5 mL LB medium containing 50 μg mL-1 ampicillin and plasmid DNA was isolated from putative recombinants using QIAprep® Spin Miniprep Kit (QIAGEN, Germany), according to the manufacturer’s protocol. The putative plasmid DNAs were sent for sequencing using T7 Promoter and SP6 universal primers.
Synthesis of SCAR primers: Based on the sequence of the cloned RAPD fragments, specific primers were designed to be used as SCAR primers. Each primer contained the original 10 bases of the RAPD primer plus another 10 bases from the flanking cloned fragment. The melting temperature, GC contents and secondary structures of each primer were verified using Gene Runner© V3.05 (http://www.generunner.net). Amplification of genomic DNA with the SCAR primers consisted of an initial hot start at 94°C, starting off with a 3 min pre-denaturation at 94°C, followed by 35 cycles of 54 sec denaturation at 94°C, 45 sec annealing at respective temperatures and 2 min extension at 72°C, with a final 5 min extension at 72°C. A series of annealing temperatures were utilized from 70 to 80°C in 1°C increments in the PCR reaction conditions in an effort to identify an annealing temperature that would yield a single and strongly amplified band of the expected size. Amplification products were electrophoretically resolved on 2.0% agarose gels with 100 bp DNA ladder (Fermentes, Canada), ethidium bromide stained and photographed under UV light using the FlourChem™ Bio-imaging System (Alpha Innotech, USA).
RESULTS
RAPD polymorphism: In this study, genetic variation was consistently detected using RAPDs in the three different Aquilaria species. RAPD analysis in 14 Aquilaria saplings yielded a total of 368 amplified reproducible bands from 23 random decamer primers Fig. 2 (a-w). The number of bands generated by a primer ranged from nine bands in OPA-20 and OPB-12, to 27 bands in OPA-18. The size of the amplified products varied from 375 to 4000 bp. All of the primers were found to be polymorphic and produced different percentages of polymorphisms.
| Table 1: | Nei’s unbiased identity and distance based on RAPD markers |
 | |
| Identity (above diagonal); Distance (below diagonal) | |
From the Multi-populations Descriptive Statistics, 333 (90.49%) polymorphic bands were found at the species level. The percentages of polymorphic loci for a single population/species were 29.08% in 107 polymorphic loci for A. malaccensis, 15.22% in 56 polymorphic loci for A. hirta and 2.99% in 11 polymorphic loci for Aquilaria sp.1.
Genetic variation: Genetic similarity was estimated within and among populations using genetic identity. Through Nei (1978) unbiased measurement of identity, all three species in this study had considerable similarities to each other. When comparing Genetic Identity (I) (Table 1) between the three species, A. malaccensis shared the highest similarity (I = 0.5780) with A. hirta. By contrast, Aquilaria sp.1 had the lowest similarity value (I = 0.5294) when compared to A. malaccensis but a relatively higher value (I = 0.5537) when compared to A. hirta. A UPGMA dendogram was constructed based on Nei’s genetic distance (Fig. 3). Similarly, the dendrogram revealed that A. malaccensis is more closely related to A. hirta while the Aquilaria sp.1 population is genetically distant from both (Fig. 3, Table 1).
Development of SCAR markers: Upon screening the 23 RAPD markers over all individuals, five distinct and reproducible bands were identified: ~950 bp from OPA02 (Fig. 2a) for A. hirta, ~635 bp from OPA05 (Fig. 2d) for Aquilaria sp.1, ~850 bp for OPA08 (Fig. 2f) for A. hirta, ~635 bp from OPA09 (Fig. 2g) for A. malaccensis and ~820 bp for OPB06 (Fig. 2o) for A. hirta. Five sets of SCAR primers were designed from the RAPD-derived sequences (Table 2). Primer specificity was evaluated using two saplings from each species. Using SCAR primer OPA05AS, amplification yielded a single distinct band for Aquilaria sp.1 at 637 bp (Fig. 4a). For A. hirta, three SCARs were developed; OPB06AH specifically amplified a band at 826 bp, OPA02AH at 955 bp and OPA08AH at 866 bp (Fig. 4b-d). In A. malaccensis, amplification using OPA09AM yielded a single band at 566 bp (Fig. 4e).
| Table 2: | Primer sequence, product length and annealing temperature of the SCAR primers for Aquilaria sp.1 (AS), A. hirta (AH) and A. malaccensis (AM) |
 | |
 | |
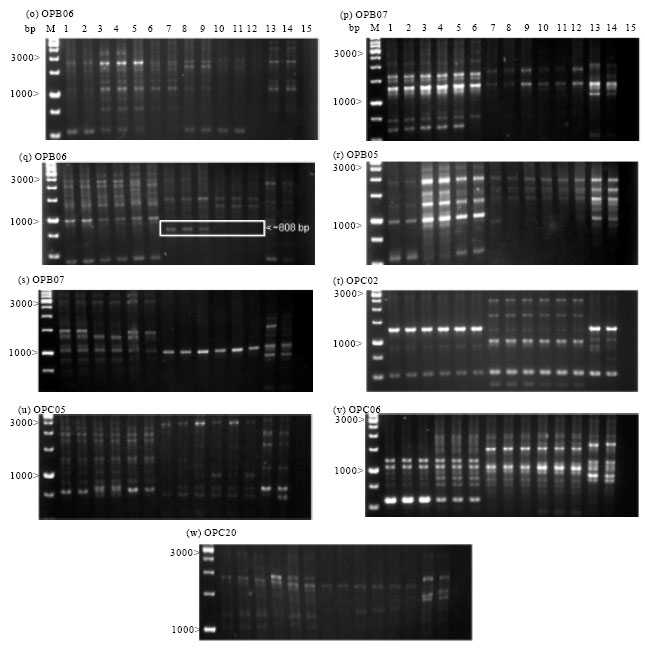 | |
| Fig. 2 (a-w): | RAPD profiles of Aquilaria species using Operon 10-mer random primers. Aquilaria malaccensis (Karak) (Lane: 1-6), A. hirta (Merchang) (Lane: 7-12), Aquilaria sp.1(Semenggoh) (Lane: 13-14) and a negative control (Lane: 15). a to w are agarose electrophoresis of the respective RAPD primers. M represents a 1 kb DNA ladder. Arrowheads and boxes indicate the species-specific RAPD amplicons used in SCAR marker development |
Specificity of SCAR marker: To examine the specificity and validity of the SCAR marker, a PCR verification test was performed on A. malaccensis from various populations. This particular species was selected because it is widely distributed in Peninsular Malaysia if compared to the other two species. Population of A. hirta is limited to a few locations in Peninsular Malaysia and many of them are inaccessible or unknown while Aquilaria sp. 1 is exclusively confined to the State of Sarawak, in East Malaysia. Genomic DNA was extracted from two individuals representing five different A. malaccensis populations including Bukit Bauk, Gua Musang, Jelebu, Jeli and Sungai Udang. Control trees representing Aquilaria sp.1 from Semenggoh, A. hirta from Merchang and A. malaccensis from Karak were also included in the testing (Fig. 1). Using OPA09AM primer in PCR, all the A. malaccensis individuals from various geographical locations yielded the expected specific band, similar to the positive control, A. malaccensis from Karak (Fig. 5).
 | |
| Fig. 3: | Dendrogram of selected Aquilaria species in Malaysia, constructed using UPGMA analysis based on Nei’s Genetic Distance calculated from 23 RAPD markers. Scale is the genetic distance measurement |
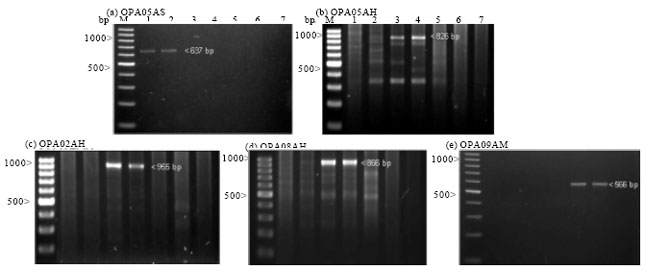 | |
| Fig. 4 (a-e): | Marker profiles of Aquilaria species generated by specific SCAR primers. Aquilaria sp.1 (Semenggoh) (Lane: 1-2), A. hirta (Merchang) (Lane: 3-4), A. malaccensis (Karak) (Lane: 5-6) and a negative control (Lane: 7). a to e are agarose electrophoresis of the respective SCAR primers. M represents a 100 bp DNA ladder. Arrowheads indicate the expected size of species-specific SCAR marker for each Aquilaria species |
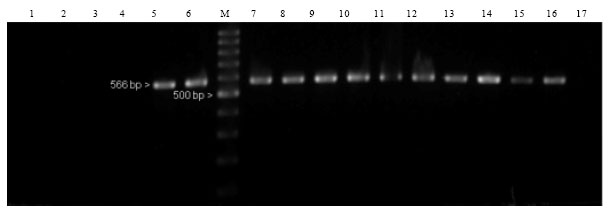 | |
| Fig. 5: | Marker profiles of Aquilaria species generated by specific SCAR primers OPA09AM developed for A. malaccensis. Aquilaria sp.1 (Semenggoh) (Lanes: 1-2), A. hirta (Merchang) (Lanes: 3-4), A. malaccensis (Karak) (Lanes: 5-6), A. malaccensis (Bukit Bauk) (Lanes: 7-8), A. malaccensis (Jelebu) (Lanes: 9-10), A. malaccensis (Jeli) (Lanes: 11-12), A. malaccensis (Gua Musang) (Lanes: 13-14), A. malaccensis (Sungai Udang) (Lanes: 15-16) and a negative control (Lane: 17). M represents a 100 bp DNA ladder. A single band of 566 bp was observed in all lanes containing A. malaccensis samples |
As expected no bands were amplified from individuals representing A. hirta and Aquilaria sp.1.
DISCUSSION
In this study, genetic variation was consistently detected using RAPDs in three Aquilaria species. Genetic identity values revealed considerable similarities among A. malaccensis, A. hirta and Aquilaria sp.1. The larger genetic distance observed between the later and A. malaccensis and A. hirta might be explained by the species geographic distributions. Sample for Aquilaria sp.1 originated from Sarawak of the East Malaysia region (Borneo Island) while samples for both A. malaccensis and A. hirta were collected from the Peninsular Malaysia region. Geographical variability is one of the common drivers that can generate high level of polymorphisms between populations, as environment differences such as the weather, air moisture content, soil nutrient, forest diversity and others can be different for both East and Peninsular Malaysia forests. Such phenomena was also supported by Josiah et al. (2008) who stated that geographical isolation may restrict gene flow, causing pollen and seed dispersal to be limited within the population and hence reduces the gene variability of the population. It is also plausible that the apparent correlation between genetic and geographic distance suggests that Aquilaria sp.1 might have been geographically isolated from A. malaccensis and A. hirta at an earlier time.
The use of RAPD permits screening through a large number of individuals in a relatively short time, providing easily-defined molecular information of a population. In addition, preliminary studies or initial works are not required before RAPD screening (Josiah et al., 2008; Sun and Wong, 2001; Williams et al., 1990). As Aquilaria had limited genetic information, RAPD allowed a quick start-off for this genus. In several cases, RAPD profiles revealed intra specific polymorphism within individuals collected in a same area. For example, within Aquilaria sp.1 from Sarawak, a clear band at 750 bp was detected in lane 14 but not in lane 13 (Fig. 2q). For identification at the species level, the dominance of RAPD markers usually does not allow to distinguish between marker/marker homozygotes and marker/null heterozygotes (Lynch and Milligan, 1994; Sunnucks, 2000). The required homozygocity of a species-specific marker can be assumed when the marker is consistently revealed for different populations. This assumption was strongly supported for A. malaccensis for which the retained RAPD and corresponding SCAR markers were consistently amplified from six samples collected in a same area and five samples collected across Peninsular Malaysia. For Aquilaria sp.1 and A. hirta, SCAR marker homozygocity and species specificity have only been tested on a reduced number of samples collected locally.
The reproducibility of RAPD results has often been questioned due to its sensitivity to parameters and the quality of the DNA template (Semagn et al., 2006). This limitation can be overcome by designing SCAR markers which, due to longer primer sequences and higher annealing temperatures are less sensitive to uncontrolled variations of PCR conditions (Jiang et al., 2009). Furthermore, in the context of quality control at the nursery, the detection of a single presence/absence signal allowed by SCARs is straight forward in comparison to the interpretation of RAPD patterns or microsatellite band lengths that may change according to intra-specific polymorphism. However, with the absence of complementary amplification patterns, false negatives may go undetected in the presence of PCR-inhibitory substances. If no false negative could be revealed during the present work, future changes in the selection of plant material and DNA extraction methods need to be thoroughly tested for their reliability before being used at the nursery. Additionally, other markers such as those based on the plastid trnL-trnF intergenic spacer and nuclear ribosomal ITS regions should be used to provide supplementary evidence in case of unexpected negative results.
In comparison to Aquilaria sp.1 and A. hirta, A. malaccensis is the most widely distributed species and is found available in all the states in Peninsular Malaysia, except Perlis. The species also supplies the bulk of agarwood traded locally and internationally. Using OPA09AM marker, a distinct band was amplified in all A. malaccensis samples collected across Malaysia but not in the other two species, leading to its possible use as control marker for fast and efficient identification of A. malaccensis. Besides being an efficient source of polymorphism for the design of species-specific SCAR, RAPD profiles have potential for use in identification of markers linked to genes or Quantitative Trait Loci (QTL) for selective breeding purposes (Collard et al., 2005; Verhaegen et al., 1997). For such application, the presence of polymorphism within the selected RAPD band sequence would allow the design of co-dominant SCAR markers (Paran and Michelmore, 1993). Yet, in the field of conservation biology, co-dominant markers such as microsatellites are usually preferred to RAPD since they are more informative for the estimation of the impact of genetic drift on genetic variation, the level of inbreeding within populations and gene flows between populations (Lynch and Milligan, 1994). Nevertheless, for a less known tropical tree like Aquilaria, RAPD and SCAR provide effective and cheap identification tool, suffice for species verification.
CONCLUSION
Previously, morphological characters have been used to describe the many different species in Aquilaria, however, species identification remained subtle as many of the characters are prone to environment conditions. In this study, RAPD markers were used for the estimation of genetic diversity in three Aquilaria species native to Malaysia. RAPD was found to be a powerful tool in assessing the genetic diversity amongst the different species and SCAR markers derived from RAPDs could serve as an efficient tool for the identification of specific Aquilaria species and could be used in breeding program.
ACKNOWLEDGMENT
This work was supported by the Universiti Putra Malaysia, Research University Grant Scheme (Project No. 03-01-09-0829RU).
REFERENCES
- Al-Saghir, M.G. and A.G. Abdel Salam, 2011. Genetic diversity of North American Soybean (Glycine max L.) cultivars as revealed by RAPD markers. J. Plant Sci., 6: 36-42.
CrossRef - CITES, 2004. Convention on international trade in endangered species of wild fauna and flora amendments to Appendices I and II of CITES. Proceedings of the 13th Meeting of the Conference of the Parties, Oct. 3-14, Bangkok, pp: 1-9.
Direct Link - Collard, B.C.Y., M.Z.Z. Jahufer, J.B. Brouwer and E.C.K. Pang, 2005. An introduction to markers, quantitative trait loci (QTL) mapping and marker-assisted selection for crop improvement: The basic concepts. Euphytica, 142: 169-196.
CrossRef - Eurlings, M.C.M. and B. Gravendeel, 2005. TrnL-trnF sequence data imply paraphyly of Aquilaria and Gyrinops (Thymelaeaceae) and provide new perspectives for agarwood identification. Plant Syst. Evol., 254: 1-12.
CrossRefDirect Link - Eurlings, M.C.M., H.H. van Beek and B. Gravendeel, 2010. Polymorphic microsatellites for forensic identification of agarwood (Aquilaria crassna). Forensic Sci. Int., 197: 30-34.
CrossRefPubMedDirect Link - Faridah-Hanum, I., M.Z. Mustapa, P. Lepun, T.I.T. Marina, M. Nazri, R. Alan and R. Mohamed, 2009. Notes on the distribution and ecology of Aquilaria Lam. (Thymelaeaceae) in Malaysia. Malaysian Forester, 72: 247-259.
Direct Link - Gokbayrak, Z. and G. Soylemezoglu, 2010. Grapevine throughout the history of Anatolia. Int. J. Bot., 6: 465-472.
CrossRef - Jena, S.N., K. Susheel and N.K. Nair, 2009. Molecular phylogeny in Indian Citrus L. (Rutaceae) inferred through PCR-RFLP and trnL-trnF sequence data of chloroplast DNA. Scientia Hortic., 119: 403-416.
CrossRef - Jiang, S., X. Jianhua and X. Li, 2009. A study on the RAPD and SCAR molecular markers of Piper species. J. Agric. Rural Dev. Trop. Subtropics, 110: 127-135.
Direct Link - Josiah, C.C., D.O. George, O.M. Eleazar and W.F. Nyamu, 2008. Genetic diversity in Kenyan populations of Acacia senegal (L.) willd revealed by combined RAPD and ISSR markers. Afr. J. Biotechnol., 7: 2333-2340.
Direct Link - Kiet, L.C., P.J.A. Kessler and M. Eurlings, 2005. A new species of aquilaria (Thymelaeaceae) from vietnam. Blumea-Biodiver. Evol. Biogeogr. Plants, 50: 135-141.
CrossRefDirect Link - Kulkarni, M. and U. Deshpande, 2006. RAPD based fingerprinting of tomato genotypes for identification of mutant and wild cherry specific markers. J. Plant Sci., 1: 192-200.
CrossRefDirect Link - Linos, A.A., P.J. Bebeli and P.J. Kaltsikes, 2002. Cultivar identification in upland cotton using RAPD markers. Aust. J. Agric. Res., 53: 637-642.
CrossRef - Lynch, M. and B.G. Milligan, 1994. Analysis of population genetic structure with RAPD markers. Mol. Ecol., 3: 91-99.
CrossRefPubMedDirect Link - Melgoza-Villagomez, C.M., H.S. Espino, A.N. Cedillo, L.L.V. Montero and C.P. Segovia et al., 2009. Identification of peach genotypes (Prunus persica (L.) Batsch) in the North-Central Region, Mexico. Int. J. Bot., 5: 160-165.
CrossRefDirect Link - Naef, R., 2011. The volatile and semi-volatile constituents of agarwood, the infected heartwood of Aquilaria species: A review. Flavour Fragr. J., 26: 73-87.
CrossRefDirect Link - Nei, M., 1978. Estimation of average heterozygosity and genetic distance from a small number of individuals. Genetics, 89: 583-590.
PubMedDirect Link - Ouborg, N.J., Y. Piquot and J.M. van Groenendael, 1999. Population genetics, molecular markers and the study of dispersal in plants. J. Ecol., 87: 551-568.
CrossRef - Paran, I. and R.W. Michelmore, 1993. Development of reliable PCR-based markers linked to downy mildew resistance genes in lettuce. Theor. Applied Genet., 85: 985-993.
CrossRef - Patra, A.P., A.K. Mukherjee and L. Acharya, 2011. Comparative study of RAPD and ISSR markers to assess the genetic diversity of betel vine (Piper betle L.) in Orissa, India. Am. J. Biochem. Mol. Biol., 1: 200-211.
CrossRefDirect Link - Ridgway, K.P., J.M. Duck and J.P. Young, 2003. Identification of roots from grass swards using PCR-RFLP and FFLP of the plastid trnL (UAA) intron. BMC Ecology, 3: 8-8.
CrossRef - Semagn, K., A. Bjornstad and M.N. Ndjiondjop, 2006. An overview of molecular markers methods for plants. Afr. J. Biotechnol., 5: 2540-2568.
Direct Link - Sun, M. and K.C. Wong, 2001. Genetic structure of three orchid species with contrasting breeding systems using RAPD and allozyme markers. Am. J. Bot., 88: 2180-2188.
Direct Link - Sunnucks, P., 2000. Efficient genetic markers for population biology. Trends Ecol. Evol., 15: 199-203.
CrossRef - Theerakulpisut, P., N. Kanawapee, D. Maensiri, S. Bunnag and P. Chantaranothai, 2008. Development of species-specific SCAR markers for identification of three medicinal species of Phyllanthus. J. Syst. Evol., 46: 614-621.
Direct Link - Verhaegen, D., C. Plomion, J.M. Gion, M. Poitel, P. Costa, and A. Kremer, 1997. Quantitative trait dissection analysis in Eucalyptus using RAPD markers: 1. Detection of QTL in interspecific progeny, stability of QTL across different ages. Theoret. Appl. Genet., 95: 597-608.
CrossRef - Williams, J.G.K., A.R. Kubelik, K.J. Livak, J.A. Rafalski and S.V. Tingey, 1990. DNA polymorphisms amplified by arbitrary primers are useful as genetic markers. Nucleic Acids Res., 18: 6531-6535.
CrossRefPubMedDirect Link - Zhang, Y.T., Z.F. Wang, H.L. Cao, X.Y. Li, L.F. Wu, S.B. Zhuo and X.F. Huang, 2010. Isolation and characterization of polymorphic microsatellite loci in Aquilaria sinensis (Lour.) Gilg. Conserv. Genet. Resour., 2: 5-6.
Direct Link