
Pradeep Kumar
Division of Crop Protection, Central Institute for Subtropical Horticulture, Lucknow, Uttar Pradesh-227107, India
Ashok Kumar Misra
Division of Crop Protection, Central Institute for Subtropical Horticulture, Lucknow, Uttar Pradesh-227107, India
Dinesh Raj Modi
Department of Biotechnology, Babashaheb Bhimrao Ambedkar University, Lucknow, Uttar Pradesh-226025, India
Asian Journal of Plant Sciences
Year: 2011 | Volume: 10 | Issue: 1 | Page No.: 1-23







ABSTRACT
Mango (Mangifera indica L.) occupies a pre-eminent place amongst fruit crops in India and is acknowledged as ‘King of fruits’ in the country. Malformation is the most threatening malady that causes great economic loss and limits the mango production in India and among tropical and subtropical countries around the globe. Floral malformation, in contrast to vegetative one, is very virulent and can cause the loss of the entire crop. Affected panicles either do not set fruit or abort fruit shortly after they have set; yields can be reduced by as much as 50-80%. Mango Malformation Disease is a fungal disease of mangoes caused by Fusarium species Fusarium moniliforme var. subglutinans. Mango is the only known host of the disease. Numerous studies on physiological, fungal, acarological, nutritional aspects have attempted, still the nature of the disorder is not fully understood. Keeping in view the seriousness of the problem, the present review explores out to establish a suitable genetic base, indicating the degree of resistance against malformation in different mango cultivars grown under open, natural field infection conditions. Disease management practices through various approaches.
PDF Abstract XML References Citation
Received: October 28, 2010;
Accepted: November 30, 2010;
Published: February 07, 2011
How to cite this article
Pradeep Kumar, Ashok Kumar Misra and Dinesh Raj Modi, 2011. Current Status of Mango Malformation in India. Asian Journal of Plant Sciences, 10: 1-23.
DOI: 10.3923/ajps.2011.1.23
URL: https://scialert.net/abstract/?doi=ajps.2011.1.23
DOI: 10.3923/ajps.2011.1.23
URL: https://scialert.net/abstract/?doi=ajps.2011.1.23
INTRODUCTION
The genus Mangifera is one among the 73 genera belonging to the family Anacardiaceae in the order Sapindales. Mango (Mangifera indica) has been
cultivated for more than 5000 years and a wide genetic diversity exist in this crop. The largest number of Mangifera species are found in the Malay Peninsula, the Indonesian archipelago, Thailand, Indo-China and the Philippines. Edible fruits are produced by at least 27 species in the genus.
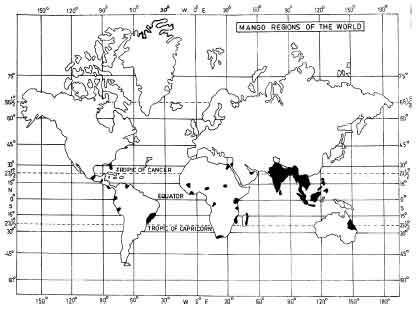 | |
| Fig. 1: | Mango growing regions of the world (Bains and Pant, 2003) |
Mangoes are native to Southern Asia, especially eastern India, Burma and Andaman Islands. It’s production predominates in dry and wet tropical low land areas 23° 26' North and South of the equator, on the Indian subcontinent, Southeast Asia and Central and South America (Litz, 1997) (Fig. 1). Mango the ‘King of Fruits’ is most popular fruit among millions of people in India and abroad. In terms of total fruit production on a global basis mango is next to banana (NBH, Indian Horticulture Database, 2008). For mango, India ranks first in area and production in the world, accounting 41.0% of the total world mango production i.e.,, 19.22 million tones(mt) (Negi, 2000). India’s contribution to the world’s mango production is highest i.e.,, 13,692,000 mt from 2,205,000 ha and about 54531 mt of mango is exported of approx. value of Rs. 127.42 crores (NBH, National Horticulture Database 2008). In spite of the highest area (22, 05,000 ha) and production of mango in the world, the mango productivity is very low i.e.,, 6.3 mt/ha. This is because of the several abiotic and biotic factors, of which malformation is one of the most important problems.
SYMPTOMOLOGY
Malformation is one of the most important problems of mango and a serious threat to mango production in India and other mango growing countries (tropical and subtropical) of the world (Crane and Campbell, 1994). It is described as disease (Summanwar, 1967; Varma et al., 1974b; Misra and Singh, 2002) and a physiological disorder (Sattar, 1946; Majumder et al., 1970a). It was first reported about hundred twenty years back from Darbhanga district in Bihar by Maries (Watt, 1891), though it has debatable issue for the scientists for the last 6-7 decades. This disorder is widespread in flowers and vegetative shoots of mango. It has a crippling effect on mango production, bringing in heavy economic losses (Hiffny et al., 1978). In spite of several decades of incessant research, since its recognition in 1891, the etiology of this disease remained confusing and no effective control measure is known (Ram and Yadav, 1999; Pant, 2000; Bains and Pant, 2003), though strong evidence is for its origin of fungal nature (Summanwar et al., 1966; Varma et al., 1974b; Misra and Singh, 2002).
Broadly three distinct types of symptoms were described by various workers. These are bunchy top of seedlings, vegetative malformation and floral malformation. Later, these were grouped under two broad categories i.e., vegetative and floral malformation (Fig. 2) (Varma, 1983a).
 | |
| Fig. 2: | Healthy and malformed inflorescence and vegetative tissue. Healthy Inflorescence. Malformed Inflorescence. Healthy Vegetative Tissue. Malformed Vegetative Tissue |
Vegetative Malformation (VM): Vegetative Malformation (VM) is more commonly found on young seedlings (Nirvan, 1953). The seedlings produce small shootlets bearing small scaly leaves with a bunch like appearance on the shoot apex. Hence, the apical dominance is lost and the seedling remains stunted and numerous vegetative buds sprout producing hypertrophied growth, which constitutes vegetative malformation. The multi-branching of shoot apex with scaly leaves is known as “Bunchy Top”, also referred to as “Witche’s Broom” (Bhatnagar and Beniwal, 1977; Kanwar and Nijjar, 1979). The seedlings, which become malformed early, remain stunted and die while, those getting infected later resume normal growth above the malformed areas (Singh et al., 1961; Kumar and Beniwal, 1992a).
Floral Malformation (FM): Floral Malformation (FM) is the malformation of panicles. The primary, secondary and tertiary rachises become short, thickened and hypertrophied. Such panicles are greener and heavier with increased crowded branching. These panicles have numerous flowers that remain unopened and are predominantly male and rarely bisexual (Singh et al., 1961; Schlosser, 1971; Hiffny et al., 1978). The ovary of malformed bisexual flowers is exceptionally enlarged and non-functional with poor pollen viability (Mallik, 1963; Shawky et al., 1980). Both healthy and malformed flowers appear on the same panicle or on the same shoot. The severity of malformation may vary on the same shoot from light to medium or heavy malformation of panicles (Varma et al., 1969a). Theheavily malformed panicles are compact and overcrowded due to larger flowers. They continue to grow and remain as black masses of dry tissue during summer but some of them continue to grow till the next season. They bear flowers after fruit set has taken place in normal panicles (Singh et al., 1961; Varma et al., 1969b; Hiffny et al., 1978; Shawky et al., 1980) and contain brownish fluid (Prasad et al., 1965; Ram and Yadav, 1999).
On the basis of compactness of panicles, malformed panicles are classified into different groups viz., heavy, medium and light (Varma et al.,1969c; Majumder and Sinha, 1972a); compact malformed panicle, elongated malformed panicle and slight malformed panicle (Rajan, 1986) and small compact type and loose type (Kumar et al., 1993). The panicles of heavy type are very compact due to excessive crowding of flowers, keep growing to form large hanging masses of flowers, most of these dry up, and hang as brown discolored bunches, but some continue to grow till the next season. The medium types of malformed panicles are slightly less compact and persist on plant for a longer time than the normal panicles. The light type is only slightly more compact than the normal panicle and does not persist on the plant. Some times, a shoot tip may bear both types of panicles i.e., healthy as well as malformed. Less frequently, a healthy panicle may contain one or more malformed branches of a few malformed flowers or vice-versa. These partially infected panicles may bear fruits up to maturity (Kumar et al., 1993).
DISTRIBUTION
World-wide distribution: Ever since the first report of its appearance at Darbhanga district (Bihar) in India by Marries (Watt, 1891), this disease has spread among the mango growing countries of the world like Australia (Peterson, 1986), Brazil (Flechtmann et al., 1970), Cuba (Padron Soroa, 1983), Egypt (Attiah, 1955), Israel and Mexico (Malo and Mc Millan, 1972; Varma et al.,1974b), Middle east (Hassan, 1944), Pakistan (Khan and Khan, 1960), South Africa (Schwartz, 1968), Sudan (Minessey et al.,1971), United Arab Emirates (Burhan, 1991), United States of America (Malo and Mc Millan, 1972; Ploetz et al., 2002) and Bangladesh (Meah and Khan, 1992). In the beginning of 2005, it is also Sultanate of Oman (Kvas et al., 2008) (Fig. 3).
India-distribution: In India, this problem is more acute in north-west regions including Jammu than the north-eastern and southern India (Chib et al., 1984; Varma et al., 1974b). In India, the disease is also reported from Maharastra and Gujrat (Burns, 1910; Burns and Prayag, 1920); Uttar Pradesh (Singh and Chakravarthi, 1935); Punjab (Singh et al.,1940); Jammu and Kashmir (Chib et al.,1984); Bihar (Mallik, 1959); Madhya Pradesh (Mishra, 2004), Himachal Pradesh (Sharma and Badiyala, 1990) and Haryana (Mehta et al.,1986). Earlier, it was reported that the region near to Hyderabad is free from this disease (Majumder and Sharma, 1990) but recently, this malady has silently entered into Andhra Pradesh (Kulkarni, 1979). Thus, mango malformation has now become a national problem (Fig. 4) affecting production in almost all of the mango growing areas of the country.
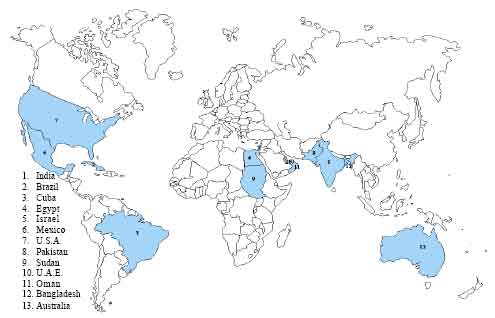 | |
| Fig. 3: | World-wide distribution of mango malformation |
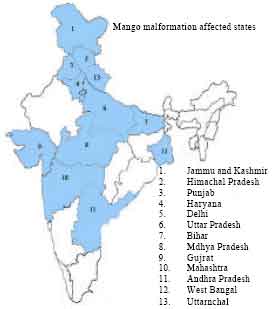 | |
| Fig. 4: | Distribution of mango malformation in India |
Malformation causes heavy damage to the trees as the inflorescence fails to produce fruits. The extent of damage varies from 50 to 60% in some cases and in severe cases the loss may be upto 100% (Summanwar, 1967; Misra et al., 2000).
CULTIVAR SUSCEPTIBILITY AND RESISTANCE
The susceptibility to malformation in mango varieties is variable and is governed by the different factors viz. temperature, age of the tree, time, etc. However, cv. Amrapalli was found highly susceptible to mango malformation particularly in Delhi regions and north India (Yadava and Singh, 1995). In general, late blooming varieties are less susceptible to malformation than the early blooming ones (Khurana and Gupta, 1973). The level of polyphenol oxidase (PPO) in the early years of plant growth or in the flush of vegetative growth may provide an estimate of synthesis of phenolic compounds in the plants, which may be correlated to susceptibility or resistance to floral malformation (Sharma et al., 1994). Based on polyphenol oxidase activity, phenolic content and panicle malformation 24 mango cultivars were classified into five groups.
The severity varies considerably from year to year. A tree once affected escapes the disease in subsequent years (Mallik, 1963). Majumder and Sinha (1972a) and Varma et al. (1969a) observed 60% diseased panicles in cv. Neelum during the flowering of February-March, whereas the same plant had only 4-5% malformation during off season flowering in June when the average minimum and maximum temperatures were higher than those of February-March, which results in decrease in disease incidence. The disease is serious in the north-west region where temperature lie from 10-15°C during December-January (winter) before flowering. The disease is mild in the areas where temperatures lie from 15-20°C, sporadic from 20-25°C and nil beyond 25°C. Fluctuations in the incidence of malformation in varieties Neelum, Alphanso and Seedling were investigated by Jagardir and Shaik (1968). Puttarudriah and Channa Basavana (1961), Singh et al. (1961) and Chadha et al. (1979a) reported that the occurrence of malformation differed according to the age of the plants. They observed more disease in young plants than in older ones. About 91% incidence in 4-8 years plants and 9.6% in older plants was reported (Singh et al., 1961). Age of the flowering shoot also influences the incidence of floral malformation, as reported by Varma (1983b).
Prakash and Misra (1986) evaluated 124 varieties of the mango germplasm against the floral malformation on the basis of %age malformed panicles. Out of these, the maximum malformation (53.14%) is recorded in variety Anopan followed by Phasla Mursidabad, Amin Prince and Bombay where disease incidence recorded is 47.37, 44.86 and 43.66%, respectively. The minimum (0-5%) incidence was recorded in 42 cvs. Misra and Prakash (1987) evaluated 274 varieties for the incidence of floral malformation in mango and found 70 varieties showing malformation 69 varieties 0-5%, 64 varieties 5-10%, 40 varieties 10-20%, 19 varieties 20-30%, 30-40% and 12 varieties showed more than 40% incidence of malformation.
Various workers in different places tried to identify resistant sources. According to Singh and Jawanda (1961) most of the mid late season varieties are more or less free from disease. Mallik (1959, 1961) found that the varieties in Bihar and UP are comparatively free from malformation. Prasad et al., (1965) reported that the seedling trees are comparatively less infested than grafted ones. Bhadauran, a monoembryonic variety was found free from malformation (Prasad et al., 1965; Majumder and Sharma, 1990; Majumder and Diware, 1985). Jawanda (1963) and Singh and Jawanda (1965) found Durre Bahisht and Kishen Bhog to be fairly tolerant. In Egypt, Azzouz et al. (1978) found cv. Zebda and Hindi Anshas were rarely affected. Misra et al. (1997, 2000) reported cv. Ellaichi as free from malformation and is now been used in breeding programme. In a recent studies cv. Meghaltan is reported disease free from malformation from West-Bengal (Ray, 2004) and cv. Dahiyar from Rewa, Madhya Pradesh (Mishra, 2004).
ETIOLOGY
The etiology of floral malformation in mango has always been controversial. However, the evidence of association of fungal pathogen(s), virus and mite has been suggested by various workers. Besides these biotic factors the abiotic factors viz. nutritional status, C/N ratio of shoots and physiological and biochemical factors for the cause of mango malformation have also been suggested from time to time by different workers.
Abiotic factors
Environmental aspects in relation to malformation: The incidence of mango malformation was found to vary from season to season but the causes of such variation are not well understood. However, studies revealed that temperature is closely related with the variation in the incidence of mango malformation (Mallik, 1963; Jagardir and Shaik, 1968).
Temperature: The seasonal variation in the temperature at the time of flowering was correlated with the intensity of malformation. The incidence of disease is most severe in north-west region of India where mean temperature during flowering remains between 10-150°C. It is mild where corresponding temperature is 15-20°C, sporadic at 20-25°C and nil over 25°C. This trend is also reflected in the world distribution of the disorder and its incidence is mainly recorded where mean temperature during winter is less than 16°C. However, the temperature dose not appear to have a major role when symptoms have already appeared on the panicles, because affected panicles continue to grow and producing excessive growth in dry hot summer (Varma et al., 1969a-c). Above 50% panicles emerging in February were found affected with malformation in cultivar Neelum, whereas only 4-5% panicles suffered in off season flowering in June. Incidence of malformation was observed to be 20% at 400 m altitude, while almost all plants were found to be free from floral malformation at an altitude of 1250 m and above. Night temperature below 10°C for long duration was found to be responsible for suppressing the incidence of floral malformation (Singh et al., 1999).
The severity varies considerably from year to year. A tree once affected escapes the disease in subsequent years (Mallik, 1963). Majumder and Sinha (1972a) and Varma et al. (1969a) studied the seasonal variation at the time of flowering with the prevailing temperature. They observed 60% diseased panicles in cv. Neelam during the flowering of February-March, whereas the same plant had only 4.5% malformation during off season flowering in June when the average minimum and maximum temperatures were higher than those of February-March which resulted in decrease in disease incidence. Fluctuations in the incidence of malformation in varieties Neelum, Alphanso and Seedling were investigated by Jagardir and Shaik (1968). Puttarudriah and Channa Basavana (1961), Singh et al. (1961) and Chadha et al. (1979a) reported that the occurrence of malformation differed according to the age of the plants. They observed more disease in young plants than in older ones. About 91% incidence in 4-8 years plants and 9.6% in older plants was reported (Singh et al., 1961). Age of the flowering shoot also influences the incidence of floral malformation, as reported by Varma (1983a).
Nutritional aspects
Micronutrient: A very little difference has been observed in the mineral constituent of healthy and malformed tissue, although micronutrient deficiency, particularly iron and zinc have been reported to be associated with the causation of malformation (Abou El-Dahab, 1975; Martin-Preveli et al., 1975; Minessey et al., 1971; Singh and Rajput, 1976). However, Pandey and Pandey, (1997b) found that zinc sulphate alone or in combination with NAA could not reduce malformation in cv. Amrapali.
Soil treatment with Bayfolan (containing N, P, K, Mn, Fe, Cu, Mg, B, Zn, Ca and Mo) @ 100 mL/tree in 22 years old trees of mango cv. Taimour also did not affect the extent of malformation (El-Beltagy et al., 1980). Malformation could not be controlled with B, Ca, Cu, Fe, Mg, Mn, K, Na, Zn or there chelates (Rajan, 1986; Ram, 1991; Saeed and Schlosser, 1972 and Tripathi, 1955). The nutrient deficiency was therefore, not considered to be a primary cause of mango malformation (Singh and Rathore, 1983).
Macro-nutrients: Reports indicate that shoot carrying malformed panicles had lower (Pandey et al., 1973, 1977) as well as higher (Mishra, 1976) levels of nitrogen than the healthy tissues. This might be due to varietal response and varying soil conditions as the former report pertains to the cv. Bombay Green under Delhi conditions and later, to the cv. Dashehari under Ludhiana conditions. Enhanced nitrogen application was found to curtail malformation, whereas addition of P and K increased the incidence significantly (Kanwar and Kahlon, 1987). However, Bindra and Bakhetia (1971) did not find any reduction in the malady by N, P and K (9:3:3) treatment and concluded that the disorder is not associated with nutritional imbalance however, these may influence the incidence of the malady. Singh et al. (1991) found lower amount of Ca in malformed tissue than in the healthy ones and suggested that calcium deficiency may not be directly responsible, although may predispose the tissue to become more prone to the incidence of this disorder. This however needs further investigations.
Metabolic imbalance (C/N ratio): The malformed tissues contain more carbohydrates and nitrogen than normal ones (Tripathi, 1955; Mallik, 1963) and higher C/N ratio was reported in malformed tissues (Khan and Khan, 1960; Pandey et al.,1973). It was postulated that high C/N ratio could be responsible for greater%age of staminate flowers on malformed panicles (Khan and Khan, 1960; Majumder and Sinha, 1972b) and suppression of development of flower and fruit set (Pandey et al., 1973). However, lower C/N ratio was observed in malformed panicles and shoots bearing them at all development stages, except for the fully swollen bud stage. This decline in carbohydrate titre and C/N ratio was attributed to enhanced utilization of these compounds and, therefore, excessive growth of malformed panicles (Singh and Dhillon, 1993b). High levels of protein were reported in malformed panicles (Tripathi and Ram, 1995). Lower rate of photosynthesis and lower leaf saccharides were observed in malformed mango cultivars (Yadava and Singh, 1995). In a similar experiment, rate of photosynthesis and chlorophyll content were low in leaves of shoots bearing clustered fruits than in leaves bearing normal fruits. However, the chlorophyll was higher in clustered fruits than normal ones. Rate of respiration, reducing sugars and activities of enzymes such as polyphenoloxidase and amylase were higher in clustered fruits (Singh and Yadav, 1997).
Physiological: Rate of photosynthesis, stomatal conductance, stomatal resistance and transpiration were found higher in healthy tissue, while rate of respiration was high in malformed tissue. The increased moisture holding capacity of malformed tissue may be due to lower transpiration (Yadava and Singh, 1995).
Biochemical aspects
Carbohydrates: There are varying reports on the status of carbohydrate in malformed and healthy panicles and shoot bearing them. The carbohydrates were invariably higher in shoots bearing malformed panicles as compared to those bearing healthy ones (Khan and Khan, 1960; Baghel et al., 1994). It was interesting to note that the higher level of starch reported in shoot bearing malformed panicles do not hydrolyze into sugar to meet their energy requirement (Pandey et al., 1977). Contrarily to this, Singh (1986b) reported the higher level of starch in healthy panicles and that of total sugar in the malformed panicles at all the developmental stages. The higher level of sugar and lower level of starch in the malformed parts than in the healthy ones suggested the hydrolysis of starch into simple sugar to meet the sufficient energy requirement for excessive growth of malformed panicles (Singh and Dhillon, 1993a).
Nucleic acids, amino acids and proteins: Free amino acids were found higher in the leaves of malformed shoots (Sandhu, 1975). In contrast, Mishra (1976) reported lower level of free amino acid in the leaves of malformed shoots in cv. Dashehari. The healthy panicles contained higher level of amino acids, RNA, DNA, soluble proteins and amides than the malformed panicles of cv. Bombay Green in Delhi and cv. Dashehri in Punjab (Pandey et al.,1976, 1977; Singh, 1986a). However this finding differed in cv. Himsagar and Bombai under West Bengal conditions, where the RNA and DNA contents increased in malformed tissue (Chattopadhyay and Nandi, 1977b, 1978). The amount of proteins and amino acids change with the stages of panicle development. The malformed panicles had more proteins and total amino acids than the healthy ones at bud inception and fully swollen stages whereas, this trend was reverse in later stages (Singh and Dhillon, 1989c-d).The total nitrogen and nitrate contents are reported higher in malformed panicles at their different developmental stages. Such type of accumulation of metabolites may be due to low catabolic activity of malformed tissue (Singh and Saini, 1997).
Phenolic compounds: The malformed panicles and shoot bearing them exhibit high level of total phenolics than the healthy ones (Mishra, 1976; and Singh and Dhillon, 1993b). However, the levels of ortho-dihydroxy phenolics in the malformed panicles and shoot bearing them varies with cultivars and agro-climatic conditions, being lower in cv. Dashehari under Punjab condition (Singh and Dhillon, 1993b) and higher in cv. Bombay Green under Delhi conditions (Pandey et al., 1977). Mangiferin (1,3,6,7-tetrahydroxy xanthone-C-β-D-glucoside), a phenolic metabolite, was either absent or present in trace in healthy panicles of Langra as against its high concentrations in malformed panicles (Ghosal et al., 1978, 1979; Chakrabarti and Ghoshal, 1989). The accumulation of mangiferin results in an excessive vegetative growth, which is subsequently transformed into the manifestation due to hormonal imbalance (Ghosal et al., 1978). Significant changes were reported in the nature and content of phenolics and steroidal constituent of healthy and malformed flowers and those artificially inoculated with the fungus Fusarium moniliforme var. subglutinans (Ghosal and Chakrabarti, 1988). Mangiferin, ellagic acid, gallic acid and a galloyl derivative of glucose were isolated from malformed and normal panicles. Two flavones, tetrahydroxy (Kaempferol) and pentrahydroxy (quercetin) were detected in the flower. There was more turnover of phenols in flowers as compared to rachis at various stages. The level of mangiferin was lower in panicles than in the vegetative shoots and leaves and the level in malformed panicles was not supraoptimal to cause abnormalities (Rajan, 1986). The phenolics and steroidal compounds may therefore, not be considered as a primary incitant of malformation, they may, however, induce toxicity symptoms but not typical malformation. These compounds may rather be the consequence of malformed growth (Kumar et al., 1993).
Enzyme activity: The higher level of IAA oxydase, peroxidase and polyphenol oxydase have been observed in malformed tissues than healthy ones (Chattopadhyay and Nandi, 1976; Beniwal et al., 1979). The higher level of glutamate dehydrogenase and lower level of nitrate reductase and glutamine synthetase were observed at early developmental stages of malformed tissue than the healthy ones (Singh et al., 1998). Rajan (1986) reported higher levels of IAA oxidase and polyphenol oxidase but lower levels of peroxidase in malformed tissue than healthy ones in cvs. Dashehari and Chausa. Pal et al. (1983) has also found higher activity of IAA oxidase but lower activity of amylase and catalase in naturally occurring as well as morphactin induced malformed panicles of cvs. Dashehari, Mallika, Benazir and Taimuria.
Roles of Plant Growth Regulators (PGRs) in the Disease Incidence
Auxins: Generally, higher level of auxins occur in healthy panicles and shoot bearing them as compared to malformed panicles and shoot bearing them. It was reported that healthy panicles as compared to malformed contained higher level of acidic, non acidic and total auxins (Dashan 1987). It was found that levels of IAA and IAN also decrease in bunchy top affected tissue by 98.4 and 92.6% respectively (Kumar et al., 1980; Dhillon and Singh, 1989; Pandey et al.,1974; Pandey, 1988). Pal et al. (1983) suggested that malformation may be due to decreased level of auxins resulting in the hormonal imbalance. Contrarily to this, higher levels of auxins have also been observed in malformed panicles (Abou-Hussein et al., 1975). The developmental stages appear to affect the nature and content of auxin in the panicles. Acidic auxins showed higher activity in malformed bud at the initial (balloon) stage but become lower than the normal ones at the rapid growth phase of panicles and increased again 32 days later. None acidic auxin, 3-indol acetonitrile (IAN) was 50 times more in malformed panicles at 48 days after bud burst (Rajan, 1986).
Cytokinins: The qualitative and quantitative changes in cytokinins of healthy and malformed panicles at different periods of their growth were studied and cytokinin concentration was found to be higher in malformed panicles than in healthy ones (Bist and Ram, 1986a). Malformed mango inflorescence contained higher levels of endogenous cytokinins than that from healthy inflorescence. The cytokinin compliment extracted from malformed flowers was different from that isolated from healthy flowers, the most striking difference being that transzeatin, dehydrozeatin and ribosyldihydrozeatin were not detected in malformed flowers and isopentenyladenine was undetectable in healthy flowers but also present in malformed flowers. The reasons for these differences were postulated to be related to the fungal infection responsible for flower malformation and could be mediated through effects of cytokinin biosynthesis and/or metabolism (Nicholson and Staden, 1988; Van Staden et al., 1989).
Gibberellins: The lower levels of gibberellins were observed in the malformed shoots than the normal ones in the susceptible cultivar Hindy in Egypt (El-Ghandour et al., 1976; Raafat et al., 1995a, b). Contrarily, there are also reports on the presence of higher levels of gibberellins in the malformed panicles in cv. Dashehari (Abou-Hussein et al., 1975; Bist and Ram, 1986a, b; Mishra and Dhillon, 1980; Singh and Dhillon, 1990a). Gibberellins activity was also found to be affected by developmental stages, location and variety. Its activity was more in normal buds from balloon to full bloom stage, which declined subsequently (Kumar and Beniwal, 1979; Rajan, 1986; Singh and Dhillon, 1990b).
A reduction in malformation of panicles was observed by the use of GA3 at the flower bud differentiation stages (Kachru et al., 1971; Shawky et al., 1978). Gibberellins like substances increased in malformed panicles and induced production of male flowers and in due course, the growth of malformed panicles increased tremendously (Abou-Hussein et al., 1975; Mishra and Dhillon, 1980; Bist and Ram, 1986b). Malformed panicles contain lower levels of gibberellins in February and higher in March-April than healthy panicles. The hormone content in malformed panicles increased sharply at full bloom in March, declined in April and reached a minimum in the first week of May, when flowering still continue in malformed panicles (Bist and Ram, 1986b). The gibberellic acid content of cultivar Dashehari was higher in malformed panicled/shoots, at bud inception (Stage II) panicles fully grown prior to full bloom (Stage III) and full grown panicle (Stage IV) except in stage I i.e, fully swollen bud stage when it was higher in healthy panicles and shoots (Singh, 1986a; Singh and Dhillon, 1986a; Singh and Dhillon, 1990a).
Ethylene: As some of the symptoms of mango malformation resemble with those of ethylene effects, several workers have implicated a role for ethylene in mango malformation (Singh and Dhillon, 1990a; Pant, 2000). Significantly higher levels of ethylene (46, 145, 67 and 34%) were detected in malformed panicles of Dashehari compared to the healthy ones at the development stages (1) fully swollen buds (2) bud inception (3) full grown panicle prior to full bloom and (iv) full grown panicle and full bloom, respectively. Similarly, shoots bearing malformed panicle show significantly higher levels of ethylene (110, 90, 106 and 104%) over those bearing healthy panicles at the four developmental stages mentioned above. Leaves borne on shoots bearing malformed panicles also had significantly more ethylene (198, 120, 79 and 159%) than the leaves on shoots bearing healthy panicles at the mentioned stages. It was suggested that the higher levels of ethylene in malformed panicles could be suppressing apical dominance of panicles, increasing isodiametric growth of rachises and thickening the secondary branches of malformed panicles producing overcrowding of flowers. The higher endogenous levels of ethylene in malformed panicles were suggested to be induced by malformin and abscisic acid (Singh and Dillon, 1990b).
Growers in India sometimes maintain smoky fires in mango orchards for several days during the vegetative flush to induce good flowering. This results in reduced malformation. This traditional practice has its own scientific merit. Smokes elevate both temperature and CO2, which in turn result in lessened ethylene production (Abeles, 1973). Vegetative malformation could be induced by smudging (Mallik, 1963). Furthermore, malformed panicles treated with 600 ppm AgNO3 were found to grow into fruit bearing healthy panicles (Bist and Ram, 1986a). The effect may certainly be due to the inhibitory effect of Ag+ in ethylene action. The success in reducing malformation by spraying 100-200 ppm naphthalene acetic acid (NAA) in the first week if October (Majumdar et al., 1970b) decidedly supraoptimal level of auxin, and the reduction of floral malformation by the spray of 400 ppm 2-Chloroethyl-PhosphonicAcid (CPA). Prior to flower bud differentiation and at bud inception may in part be due to auto inhibition by ethylene (Riov and Yang, 1982). All these reports directly/indirectly propose a role for ethylene in malformation.
Abscisic acid: Inhibitory activity of ABA was reported more in malformed panicles rather than in healthy panicles (Pandey et al., 1974; Mishra and Dhillon, 1978; Rajan, 1986; Singh and Dhillon, 1990a). The malformed panicles were treated with different concentrations of ABA (100-500 ppm) at the end of February during early stages of panicle growth. From the data it was postulated that ABA treatment decreased panicle length and girth without any effect on sex ratio. However, 220 ppm ABA brought early anthesis in the malformed panicles and 200-300 ppm ABA decreased some external symptoms of malformation in the panicles. However, higher concentration of ABA (400-500 ppm) did not show such an effect and none of the ABA treated panicles fruited (Raina and Ram, 1991).
Hydrogen cyanide: Cyanide derived from ethylene biosynthesis could contribute to the development of tissue necrosis during hypersensitive response of plants (Siefert et al., 1995, Grossman, 1996). Increased levels of cyanide due to ‘stress etylene’ may result in the accumulation of toxic level of cyanide resulting in the necrosis and death of malformed tissue of mango (Kukreja and Pant, 2000). Hydrogen cyanide applied as Dormex increased the incidence of floral malformation, suggesting therefore, an involvement of hydrogen cyanamide or like substances in causing malformation (Pandey, 1991, 1993; Pandey and Pandey, 1997a).
Other Factors Responsible for Mango Malformation
Malformin: The presence of malformin-like substances in fully grown malformed panicles has been reported (Pandey and Ram, 1994; Ram and Bist, 1984; Singh and Dhillon, 1987a). The stem and root of malformed vegetative seedlings contain a high level of malformin-like-substances and other malformins as against none in healthy seedlings (Raina and Ram, 1991; Singh, 1986b Singh and Dhillon, 1987a). Malformin like-substances some how appear to be involved in causation of malformation, with the malformin-stimulating ethylene production, manifest into hormonal imbalance, consequently disturb metabolism and induce malformation (Singh and Dhillon, 1987b). Application of antimalformins like glutathione, ascorbic acid and silver nitrate caused disappearance of malformin from panicles which fruited like healthy controls. Malformins may cause imbalance of growth substances and conditioning of host cells to produce malformed growth (Bist and Ram, 1986a; Singh and Dhillon, 1987a, 1989d, Raina and Ram 1991). Fusarium moniliforme var. intermedium (Pandey and Ram, 1994) and F. moniliforme var. subglutinans and F. moniliforme Sheld (Kumar and Ram, 1998) secreted malformins into their culture filtrates which were similar to mango malformins. Mango malformin treated mango seeds and branches of bearing trees produced malformed seedlings and shoots (Tripathi and Ram, 1998). The exact mechanism of ethylene synthesis in plant system, its chemical nature and whether ethylene production is a cause or consequence of malformation are yet to be fully worked out.
Mangiferin: Mangiferin, a non-toxic polyphenol and normal metabolite in mango was reported to play a significant role in the disease (Ghosal et al., 1979). Mangiferin is either absent or is present only in traces in healthy mango panicles but in malformed ones its concentration is high (Chakrabarti and Ghosal, 1989). Enhanced production of mangiferin and increase in the activity of polyphenol ozidase in infected tissues was recorded. Poliphenol oxidase was considered as mangiferin degrading enzyme (Kumar and Chakrabarti, 1992).
BIOTIC FACTORS
Different biotic factors are reported to be as causal agent of mango malformation disease which is directly or indirectly responsible to spread disease incidence. These factors include viruses, mites and fungus.
Viruses: Sattar (1946) considered the disease either of viral nature or a physiological disorder. Viral disease like symptoms of the malady and failure to isolate any pathogenic organism led to speculation that the disease is of viral nature (Singh and Jawanda, 1965; Giani, 1965). Mallik (1963) transmitted the disease successfully by grafting or budding, and also by dodder, but no precaution was taken to prevent the movement of mites in this study. In his studies, virus was considered to be the main cause of the disease just after the failure of physiological theory, but it soon started loosing ground, when Singh et al. (1961) and Prasad et al. (1965) tried to transmit the disease from branches to seedlings, seedlings to scions and seedlings to seedlings by anarching, cleft grafting, bark patch budding, mechanical inoculation or through insects without any success. Viral and mycoplasma like nature could not be confirmed by even Beniwal and Bhatnagar (1975). The disease does not appear to be transmitted through stones (Kausar, 1959; Bindra and Bakhetia, 1971). The results thus obtained from these studies do not suggest that mango malformation is of viral etiology (Varma, 1983b).
Mites (Acarobiogical): Association of mites with malformed panicles came into picture in 1944 when Hassan (1944) and Sayed (1946) reported the mite Aceria mangiferae (Sayed) syn. Eriophyes mangiferae from Egypt. Sayed (1946) observed the association of this mite to cause both vegetative and floral malformation. In India, Narasimhan (1954) for the first time claimed eriophid mite to be the disease causing organism and found it inter and intracellularly in the meristem and tender regions (Narasimhan, 1959). Mites were found to induce the disease in test plants when transfered from diseased (Singh et al., 1961) or even from healthy plants. (Puttarudriah and Channa Basavanna, 1961; Nariani and Seth, 1962) successfully induced the disease on young seedlings by introduction of eriophyid mites but it was not certain whether these mites were vectors of some pathogen or is a direct cause. Narayanan and Ghai (1961, 1963) and Malo and Mc Millan (1972) also found association of mite Aceria mangiferae Sayed on malformed panicles. Ahmad and Anwar (1965) were also successful in reproducing the disease through mites. Besides, A. mangiferae, Singh (1955, 1957) reported two more species of mites, namely, Typhlodromus castellanii Hirst and Typhlodromus sp. (probably asiaticus) causing malformation. Since Typhlodromus spp. are predatory mites, these can not be associated with malformation. Narayanan and Ghai (1961, 1963) recorded A. mangiferae and four other predatory mites, viz., Typhlodromus rosanlali, T. rhenamus, T. nesbitti and Chaloetogenes ornatus and were of the opinion that because of these predaceous mites, the population of the phytophagous mite (A. mangiferae) do not rise high.
Prasad et al. (1965) advocated that there is no correlation of mite and bud malformation and thus to presume that mite is the only causal organism is doubtful. Prasad et al. (1972) also found the presence of mites in buds when the scales were a bit loose indicating that mites are not the causative organism in malformation malady. Sternlicht and Goldenberg (1976) advocated that A. mangiferae has no direct effect on inflorescence malformation, but it’s interaction with another biotic factor (a fungus) is not ruled out. In Egypt, Wabha et al. (1986) suggested that A. mangiferae does not cause the malady directly but may play a role in the overall malformation process. Labuschang et al. (1993) could not correlate that presence of mites and increase in the incidence of malformation. However, role of mite as a carrier of fungus (Fusarium monilifome var. sublgutinans) was advocated by Summanwar (1967), Summanwar and Raychoudhury (1968) and Pinkas and Gazet (1992).
Fungus: Summanwar et al., (1966) reported for the first time, a fungus Fusarium moniliforme Sheld, associated with malformation (floral and vegetative) and proved its pathogenicity. Summanwar and Raychaudhuri (1968) investigated that mite carried the fungus on the bodies and irritation caused by the mites paved the way for the fungus. The fungus, F. moniliforme, has been consistently isolated from various parts of affected malformed plants (Varma et al., 1969a, 1969b; 1971, 1972, 1974a, b). Isolations were made from malformed panicles and of 392 isolates from 130 trees, 336 were Fusarium moniliforme and 36 Cylindrocarpon mangiferum (Chowdhary and Varma, 1986). The fungus was further identified as F. moniliforme var. subglutinans (Varma et al., 1974b). Aerial mycelium appearing powdery due to microconidia which are 0-1 septate, oval to fusiform and are produced on polyphialides, macroconidia are lacking or rarely produced, 1-2 septate, falcate and without chlamydospores. Pigmentation is typical violate. The fungus does not have any special nutritional requirement as reported by Chattopadhyay and Nandi (1981). Varma et al. (1971) reported that the growth of the fungus was inhibited during summer months even at room temperature. Vegetative malformation (Prasad and Singh, 1972; Summanwar et al., 1966; Varma et al., 1969a) and floral malformation (Varma et al.,1974a, b) can be initiated in the healthy test plants by artificial inoculation of aerial branches with the fungus as it is mostly intercellular and occasionally forms intercellular agglomerates in the cortex and phloem regions and the fungus form globose bodies similar to chlamydospores, particularly in the cortex when inoculated with spore suspension (Varma et al., 1972; 1974b). Cross inoculation studies with strains of F. moniliforme further confirmed the host specificity and the mango strains only caused typical disease symptoms and infection (Varma et al., 1974a, b). Definite evidence (Ghosal et al., 1977) has revealed that the disease is caused by F. moniliforme var. subglutinans. Fusarium pathogens in contact with the other host species produce the common toxic compounds namely fusaric acid, lycomarasmin and 12-13 epoxytrichothecences.
Typical bunchy top symptoms can be produced in seedling by inoculating the fungus through soil. The fungus is systemically present in parenchymatous cells of the pith region of malformed tissues (Bhatnagar and Beniwal, 1977). There is possibility that the disease is caused by more than one species of Fusarium.
The role of the Fusarium subglutinans in mango malformation has recently been emphasized again. Typically symptoms of mango malformation such as short internodes, stubby leaves and bunchy inflorescence were reported when healthy mango plants were inoculated with GUS transformants of Fusarium subglutinans. Infected plants organs contained the GUS-sustained mycilium of the pathogen (Freeman et al., 1999, 2000).
Fungal etiology and behaviour of mango malformation was also studied by Andotra et al. (1984). Inoculation of a Fusarium spp. in mango seedlings showed that the disease is neither systemic, nor completely localized but behaved erratically. Internal spread is always acropetal and is supposed to be facilitated through active cell divisions of terminal growth under environmental conditions favourable to both host and pathogen.
Effect of F. moniliforme var. subglutinans infection on mangiferin production in the twigs of Mangifera indica was studied by Chakrabarti and Ghoshal, (1985). Infected twigs contain less mangiferin than twigs of healthy plants. In both cases, mangiferin concentration was high during cooler months and low during hotter months.
Misra and Singh (1998, 2002) did studies on mass isolation and recovered above 82% F. moniliforme var. subglutinans colonies. They also isolated Fms from normal apical shoot and healthy panicles and advocated its systemic nature. Isolations made from the different portions of the normal panicles revealed highest number of Fms colonies from the base portion (69.4%) indicating the accumulation of pathogen at certain portions and threshold level of pathogen at the base portion may be responsible for the production of malformation symptom in the mango plants. Maximum Fms colonies were recovered from the samples of second fortnight of February, indicating its maximum activity during that period. Inoculation during December by breaking swollen buds and placing Fms give better success in symptom production.
The role of Fusarium subglutinans in mango malformation has recently been emphasized again. Typically, symptoms of mango malformation such as short internodes, stubby leaves and bunchy inflorescence were reported when healthy mango plants were inoculated with GUS transformants of Fusarium subglutinans. Infected plants organs contained the GUS-stained mycelium of the pathogen (Freeman et al., 1999, 2000).
Youssef et al. (2007) sampled vegetatively malformed mango seedlings, growing under infected trees bearing infected panicles in two locations in Egypt to determine whether infection in seedlings was systemic (evenly distributed within plant tissue) or whether the pathogen originated from malformed panicles. According to PCR-specific primer amplification, the pathogen was detected in 97% of seedling apical meristems, declining gradually to 5% colonization in roots. It was concluded that inoculum of the pathogen originates from infected panicles and affects seedlings from the meristem, with infections descending to lower stem sections and roots. Minor infections of roots may occur from inoculum originating from infected panicles, but the pathogen is not seedborne.
Conditions affecting germination and growth of Fusarium mangiferae, causal agent of mango malformation disease, were studied in vitro by Atinksky, et al. (2009a). Both conidial germination and colony growth required temperatures >5°C and reached a peak at 28 and 25°C, respectively. A minimum 2-h wetness period was required for conidial germination, reaching a peak after 8 h of wetness. High incidence of fungal colonization in buds, predominantly the apical buds, was detected compared with inoculated leaves. Dry, malformed inflorescence debris serving as a source of inoculum caused significantly higher colonization (52-20%) of inoculated buds, compared with that (0%) of the untreated controls. Incidence of sampled leaf disks bearing propagules of F. mangiferae from an infected orchard peaked in June and July and decreased during the following months, whereas airborne infections on 1-month-old branches was the highest in May and June, corresponding with inoculum availability released from infected inflorescences. Colonization pattern, determined in naturally infected vegetative and woody branches, was significantly higher in node sections than in the internode sections. This study sheds light on infection dynamics, colonization patters and the disease cycle of F. mangiferae in mango.
Atinksky et al. (2009b) advocated that inoculum availability and conidial dispersal patterns of Fusarium mangiferae, causal agent of mango malformation disease, were studied during (2006-2007) in an experimental orchard. The spatial pattern of primary infections in a heavily infected commercial mango orchard corresponded with a typical dispersal pattern caused by airborne propagules. Malformed inflorescences were first observed in mid-March, gradually increased, reaching a peak in May, and declined to negligible levels in August. The sporulation capacity of the malformed inflorescences was evaluated during three consecutive months. Significantly higher numbers of conidia per gram of malformed inflorescence were detected in May and June than in April. Annual conidial dissemination patterns were evaluated by active and passive trapping of conidia. A peak in trapped airborne conidia was detected in May and June for both years. The daily pattern of conidial dispersal was not associated with a specifically discernable time of day, and an exponential correlation was determined between mean Relative Humidity (RH) and mean number of trapped conidia. Higher numbers of conidia were trapped when RH values were low (<55%).
The role of the mango bud mite, Aceria mangiferae, in carrying conidia of Fusarium mangiferae, vectoring them into potential infection sites and assisting fungal infection and dissemination was further studied by Gamliel-Atinksky et al. (2009). Following the mite’s exposure to a green fluorescent protein-marked isolate, conidia were observed clinging to the mite’s body. Agar plugs bearing either bud mites or the pathogen were placed on leaves near the apical buds of potted mango plants. Conidia were found in bud bracts only when both mites and conidia were co-inoculated on the plant, demonstrating that the mite vectored the conidia into the apical bud. Potted mango plants were inoculated with conidia in the presence or absence of mites. Frequency and severity of infected buds were significantly higher in the presence of mites, revealing their significant role in the fungal infection process. Conidia and mite presence were monitored with traps in a diseased orchard over a 2-year period. No windborne bud mites bearing conidia were found; however, high numbers of windborne conidia were detected in the traps. These results suggest that A. mangiferae can carry and vector conidia between buds and assist in fungal penetration but does not play a role in the aerial dissemination of conidia between trees.
Molecular characterization of Fusarium moniliforme var. subglutinans
A mango malformation pathogen: Cultural and morphological studies give an idea about the variability in the pathogen but do not give definite answer. Studies of pathogenic variability in perennial plants have its own limitations. Recent studies on genetic variability using DNA based markers in these pathogens have opened new avenues to use molecular markers as a fast and accurate method for diagnosis of the pathogen and to differentiate and characterize different isolates (pathogenic and Vegetative Compatibility Groups (VCG’s)) and identify distinct races of the pathogens. These novel techniques are also useful in mapping of the genetic diversity occurring in the pathogens. The consolidated information on cultural, morphological, pathogenic variability and molecular characterization can be put to effective use in identification of definite distinctive strains amongst the pathogen, which can be used to understand the mechanism of resistance and subsequently streamlining the resistance breeding programme.
A molecular marker is a DNA sequence that is readily detected and whose inheritance can be easily be monitored. The uses of molecular markers are based on the naturally occurring DNA polymorphism, which forms basis for designing strategies to exploit for applied purposes. A marker must to be polymorphic i.e., it must exit in different forms so that chromosome carrying the mutant genes can be distinguished from the chromosomes with the normal gene by a marker it also carries. Genetic polymorphism is defined as the simultaneous occurrence of a trait in the same population of two discontinuous variants or genotypes. DNA markers seem to be the best candidates for efficient evaluation and selection of plant material. Unlike protein markers, DNA markers segregate as single genes and they are not affected by the environment. DNA is easily extracted from plant materials and its analysis can be cost and labour effective. The first such DNA markers to be utilized were fragments produced by restriction digestion-the Restriction Fragment Length Polymorphism (RFLP) based genes marker. Consequently, several markers system has been developed.
In recent years, numerous DNA based fingerprinting methods that reveal the genetic diversity or pathogenicity of similar organisms has been developed. RAPD and RFLP analysis have provided more rapid methods in a wide variety of species. PCR based method of gene typing based on genomic polymorphism is a recent method. This is a technique widely used for assessment of inter and intra specific genetic variation (Williams et al., 1990; Welsh and McClelland, 1990). RAPD analysis has been applied widely in the detection and gene characterization of phytopathogenic fungi (Miller, 1996; Miller and Conn, 1980).
RAPD technology in Genetic diversity analysis of Fusarium spp: RAPD is the most simplest and promising tool for genetic diversity. Despite the limitations of dominance and non-reproducibility, but for the reason of large volume of data generated, this marker has been considered as an efficient tool for genetic diversity studies, gene tagging for disease resistance through either Bulk segregant analysis or selective genotyping. The RAPD technology has quickly gained widespread acceptance and application allowed for the rapid generation of reliable, reproducible DNA fragments or fingerprints in a wide variety of species, including several fungi (Crowhurst et al., 1991; Goodwin and Annis, 1991; Khush et al., 1992). Moreover, it has been extensively used as an auxiliary tool for the genetic diversity analysis, classification, or identification of soil-borne pathogens such as Fusarium (Angela et al., 1996; Choi et al., 1997, Kini et al., 2002;, Saharan et al., 2006; Gupta, 2009) Rhizoctonia (Lee et al., 1998), or Colletotrichum (Vakalounakis and Fragkiakakis, 1999, Gupta et al., 2010). Different molecular markers like RFLP (Kelly et al., 1994, Bogale et al, 2007), RAPD (Grajal-Martin et al., 1993; Iqbal et al., 2006; Gupta et al., 2009), AFLP (Abd-Elsalam et al., 2003; Kiprop et al., 2002; Sivramakrisnan et al., 2002) had been investigated for the molecular characterization of Fusarium. RAPD has proved successful in generating amplification patterns specific to Fusarium mangiferae (Ploetz et al., 2002; Iqbal et al., 2006) and Fusarium spp. (Gupta et al., 2009).
In recent years, DNA polymorphisms have increasingly been used to complement traditional markers for the analysis of genetic identity, variability and relatedness in fungi (Kaemmer et al., 1992; Jamil et al., 2000). Molecular markers reveal information concerning the genetic structure of pathogen populations. DNA fingerprinting has evolved as a major tool in fungal characterization. Attempts to characterize fungal isolates through Random Amplified Polymorphic DNA (RAPD) have been reported (Williams et al., 1990; Grajal-Martin et al., 1993). The development of RAPD has allowed the rapid generation of reliable, reproducible DNA fragments or fingerprints in wide variety of species, including several fungi (Crowhurst et al., 1991; Goodwin and Annis, 1991; Khush et al., 1992). It has proved successful in generating amplification patterns specific to F. mangiferae (Ploetz et al., 2002).
Zheng and Ploetz (2002) examined genetic diversity among seventy four F. subglutinans isolates collected from Brazil, Egypt, Florida (USA), India, Israel and South Africa. A pair of 20-mer primers developed from a RAPD fragment amplified a specific 608 bp fragment for 51 out of 54 mango isolates. Ploetz et al. (2002) amplified genomic DNA of F. mangiferae isolates with 33 arbitrary decamer primers. Most of the primers produced reproducible polymorphic banding patterns. Unique Major bands were generated with primers OPZ-5 and OPJ-l. RAPD analysis divided the isolates into two genetically distinct clusters.
Saleem (2004) identified twenty isolates of F. mangiferae from malformed tissues of mango obtained from different areas of Pakistan. No site specific correlation was found. The potential of RAPD to identify DNA markers related to intraspecific diversification of the pathogens led to study the genetic diversity within F. mangiferae population. KaleWright et al. (1992) classified isolates of Fusarium sp. collected during a large scale survey in Victoria, by cultural morphology, pathogenicity and a RAPD assay. Morphological and cultural characteristics did not correlate with the other techniques used and were not consistent within groups.
Jimenez et al. (2001) examined twenty nine isolates of Fusarium spp. from different geographical regions for genetic relatedness using RAPD. The technique permitted to find interspecific differences among the isolates of Fusarium spp. from different hosts by combining morphological approaches with molecular analysis. Mitter et al. (2002) analyzed sixteen strains of Fusarium spp. from different geographical regions and hosts for genetic relatedness by RAPD. The analysis showed completely different pattern between high, moderate and low gibberellic acid producing strains. High producers formednearly identical RAPD patterns, whereas the low and moderate producers gave heterologous amplification patterns. These investigations found an application in distinguishing isolates of Fusarium spp. within the section Liseola. Jana et al. (2003) studied genetic variation in 22 isolates of Fusarium species collected from geographically distinct regions over a range of hosts, using RAPD markers. UPGMA analysis classified these isolates into five major groups using primer OPA-l3. Isolates of pathogenic and non-pathogenic Fusarium species of different formae-speciales and races were distinguished among each other. A robust and repeatable PCR-assay was developed for the detection and differentiation of F. sporotrichiodes from other Fusarium species based on sequence determined from differentially amplified RAPD-PCR products. These assays were able to detect both species in samples of grain taken from the field (Wilson et al., 2004).
NATURAL SPREAD OF DISEASE
The annual recurrence of malformation in new seedlings clearly shows its natural spread. Increase in infection has also been reported by Nirvan (1953), Singh et al. (1961) and Mallik (1963). Although the fungus F. mangiferae does not sporulate in situ, it does soon drying malformed panicles (Varma et al., 1974a). To study the aerial flight of these fungal spores, the use of rotary traps for six months in places having high incidence of mango malformation could not yield the spores of Fusarium (Varma et al., 1971), which clearly indicates the role of some other agency in transmission of the disease. Puttarudriah and Channabasavana (1961), Singh et al. (1961) and Nariani and Seth (1962) successfully reproduced the disease by transferring the mites which have been later reported to carry the fungus F. moniliforme on their body surface (Summanwar and Raychaudhari, 1968). Varma et al. (1971) explained the possibility of mites in transmission of disease. This finding was supported by the feeding behavior of the mites (Varma, 1983a). The small%age of mites carrying the fungus and their presence in south-eastern parts of India (Varma et al., 1971), where the disease is sporadic in nature, also indicate the possibility of some additional factor(s) in the spread of the disease (Varma et al., 1974a, b). The propagation and distribution of diseased plant material may cause wide and erratic distribution of the disease (Varma et al., 1971).
MANAGEMENT
Along with the attempts to determine the etiology of mango malformation numerous control measures have been tried but the results are neither successful nor reproducible. Various approaches to control the disease are discussed below:
Disease management through plant Growth regulators: Reduction in the malformation of panicles was observed by the use of GA3 at flower bud differentiation stage (Kachru et al., 1971; Shawky et al., 1978). Further it was reported that 200 or 500 ppm chlormequat mixed with 100 mL Bayfolan treatment reduced%age of malformed panicles (El-Beltagy et al., 1980). The application of NAA, 100-200 ppm, at flower bud differentiation stage reduced malformation considerably (Majumder et al., 1970a; Shant, 1975; Singh et al., 1977; Bajpai and Shukla, 1978; Singh and Dillon, 1986b). Spraying 200 ppm NAA in first week of October followed by sparaying of 500 ppm etherel at bud inception stage during February was highly effective in reducing floral malformation (Singh and Dhillon, 1986a).
Treatment with anti malformins like glutathione (560 ppm), ascorbic acid (1055 ppm), K2S2O5 (560 ppm), silver nitrate (2400 ppm) and NAA (200 ppm) resulted in 87, 93, 80 and 40% conversions to healthy panicles respectively (Ram and Bist,1984). Substantial reduction in floral malformation by antimalformin spray at but inception stage was observed (Singh and Dhillon, 1989a). Sparaying 1000 ppm paclobutrazol (10-60 g-1tree), prior to FBD, during the first week of October, reduced malformation, increased number of healthy flowers and increased yield (Singh and Dhillon, 1989b).
Disease management through deblossoming: Deblossoming at the bud burst stage alone or in combination with spraying of 200 ppm NAA was reported to be very effective in controlling malformation. Deblossoming at bud burst stage gives substantial reduction in malformation (Singh et al., 1974; Majumder et al., 1976; Singh et al.,1979; Singh and Dhillon, 1986b, 1988). Deblossoming between 20th January and 25th February regenerated new panicles in the same season. Regenerated panicles bore fruits similar to healthy once. Deblossoming after February failed to regenerate panicles (Tripathi and Ram, 1998).
Since, the process of deblossoming is cumbersome it is advisable to develop a chemical for deblossoming. Application of 200 and 500 ppm etherel completely control malformation (Chadha et al., 1979b). 250 ppm of cycloheximide was also very effective in deblossoming the panicles (Pal and Chadha, 1982). 750 to 6000 ppm dikegulac and 500 ppm etherel at bud burst stage were ineffective (Singh and Dhillon 1986a).
Disease management through nutrients: In some cases, nutrient application has improved the nutritional statusthus improving yield. A direct inhibitory effect of chemicals against pathogen is inferred together with secondary control through improved nutritional status of trees. The combined effect of potassium sulphate as soil application and monocrotophos as trunk injection cured the malady. Result of a ten year trial of NPK fertilization on panicle malformation in mango cv. Dashehari indicated that increasing nitrogen doses reduced panicle malformation whereas the effect of phosphorus and potassium was just the reverse (Minessey et al., 1971; Abou-EL-Dahab, 1975). Partial control of the disease has been achieved in India by spraying the malformed parts with mangiferin-Zn2+ and mangiferin-Cu2+ chilates (Chakrabarti and Ghosal, 1989).
Disease management through pruning of malformed parts: Pruning reduced malformation (Narasimhan, 1959; Desai et al., 1962; Singh et al., 1983). Pruning however, had no effect on malformation (Bindra and Bakhetia, 1971). Pruning followed by spraying with the mixture of fungicide (Captan 0.1%), miticide (Akar 338-0.1%) and Sticker (Tenae) helped considerably in controlling at least the spread of the disease (Summanwar, 1967). Pruning of diseased parts and spraying with diazinon were reported to control the malady (Rai and Singh, 1967; Yadav, 1972).
Disease management through insecticides and fungicides: Pruning followed by a spray of insecticides viz., follidol and /or metasystox as a control measure was recommended (Giani, 1965). Aceria mangiferae was effectively controlled by a spray of 0.15% phosphon or Formothion (Wafa and Osman, 1972). Significant reduction in the malady was reported by applying various insecticides (Giani, 1965; Diekmann et al., 1982).
Fungal theory necessitated the use of systemic fungicides (Varma et al., 1971). It was concluded that copper fungicides were superior to organic fungicides due to excellent tenacity under monsoon conditions. The inhibitory effect of different fungicides viz., Fytolan, hexaferb and captan on mango plants artificially inoculated with F. moniliforme var. subglutinans was studied (Chattopadhyay and Nandi, 1977a). Fytolan was maximally effective at all concentration. In view of experimental evidences and economy of fytolan, it may be recommended for controlling malformation of mango in areas where the disease is a serious problem (Chattopadhyay and Nandi, 1978). Benomyl failed to control the problem in South Africa (Diekmann et al., 1982) and in India but some success in reducing disease severity by spray application of benomyl has been reported in India (Siddiqui et al.,1987) and in Israel (Pinkas and Gazit, 1992). Carbendazim through trunk injection or soil application, either alone or in combination with cultural practices such as root pruning showed no improvement (Kumar and Beniwal, 1992b).
In in vitro, evaluation Carbendazim was found to check the growth of F. subglutinans completely at 0.1%. Hence, a trial was laid out for the control of malformation with spray of Carbendazim (Bavistin 50 W.P.) 0.1% at 10, 15 and 30 days interval starting from October to February (consisting of 13, 9 and 5 sprays, respectively) during the flower bud differentiation stage. It was revealed from the data, that malformation incidence was least in 10 and 15 days interval spray interval schedule and between the two spray schedules, there was no difference. Control of the malformation was up to 76.93% in 10 and 15 days spray schedule, while it was 65.39% in 30 days spray schedule over check. Thus, fifteen days interval spray schedule is recommended for the control of the disease (Misra et al., 2000).
Disease management through biopesticides: Three different species of Trichoderma i.e., Trichoderma viride, T. virens and T. harzianum were tested against the F. moniliforme var. subglutinans (Kumar et al., 2007). The three bioagents varied in their efficacy against F. moniliforme var. subglutinans isolates. In general, all the three bioagents were effective in checking the growth of all evaluated isolates of Fusarium. However, out of the three bioagents best result was obtained with T. harzianum followed by T. virens and T. viride. Results clearly showed that the% inhibition of Fusarium isolates by T. harzianum was significantly superior to T. viride for all the isolates.
Kumar et al. (2009) further evaluated leaf extract of twenty-three plants for their antifungal activity against F. moniliforme var. subglutinans. Although, all the leaf extracts checked the radial growth of test fungus, extracts of Azadirachta indica, A. Juss., Achyrenthes roseus and Calotropis gigantea were found more effective against F. subglutinans. However, the leaf extract from Aegle marmelos (L.) Corr., Ricinus communis L. and Ficus racemosa L. were found less effective. Usha et al. (2009) also reported antifungal activity of Dhatura stramonium, Calotropis gigantean and Azadirachta indica against floral malformation pathogens.
CONCLUSION AND FUTURE PROSPECT
Fusarium is one of the most heterogeneous fungal genera and classification of species within this genus is very difficult. Currently, the differentiation of Fusarium spp. is based on physiological and morphological characteristics such as the shape and size of the macroconidia, the presence or absence of microconidia and chlamydospores, and colony morphology. Subtle differences in a single characteristic may delineate species. These observations need some practice and are difficult for the non-specialist. F. moniliforme var. subglutinans is the causal agent of malformation in mango (Mangifera indica L.) in the form of malformation on leaves, apical shoots, flowers and inflorescence. Accordingly, the malformation on former two portions is described as vegetative malformation and the latter two portions as floral malformation. Infection on flowers results in severe losses. The symptoms on shoots and flowers are not always identical and sometimes may be attributed to environmental conditions or existence of variability among the pathogenic isolates from different agro-climatic regions. The morphological and physiological methods used for identification and classification of F. moniliforme isolates have proved problematic. Variability, taxonomic and evolutionary studies in fungi have increased with the development of molecular techniques. PCR-based fingerprinting techniques (RAPDs, SSRs and AFLPs) offer highly sensitive, easy, rapid detection method and less time consuming for diagnosis of pathogen. Random Amplified Polymorphic DNA (RAPD) is a DNA polymorphism assay based on the amplification of random DNA segments with single primers of arbitrary nucleotide sequence. Therefore, a rapid and reliable assay for the detection of F. mangiferae var. subglutinans would benefit the mango production. Molecular marker based characterization of mango malformation pathogen using RAPD markers for genetic diversity analysis and development of a SCAR marker based diagnostic kit would be a possible solution to early detection and diagnosis of mango malformation disease. In addition to contributing to the understanding of the diseases caused by this pathogen and improving crop productivity, this information will improve the basic understanding of the molecular biology of this pathogen and will be of importance to many other agricultural systems.
REFERENCES
- Abd-Elsalam, K.A., F. Schnieder, M.S. Khalil, A. Asran-Amal and J.A. Verreet, 2003. Use of AFLP fingerprinting to analyze genetic variation within and between populations of Fusarium spp. derived from Egyptian cotton cultivars. J. Plant Pathol., 85: 99-103.
Direct Link - Andotra, P.S., H.S. Chib and B.R. Gupta, 1984. Studies on the fungal etiology and behaviour of mango malformation under Jammu conditions. Indian J. Mycol. Plant Pathol., 14: 90-92.
Direct Link - Angela, G.S., M.M. Evelyn and H.G. Hartwig, 1996. Polymerase chain reaction-based assays for species-specific detection of Fusarium culmorum, F. graminearum and F. avenaceum. Phytopathology, 86: 515-522.
Direct Link - Atinksky, E.G., S. Freeman, A. Sztejnberg, M. Maymon, R. Ochoa, E. Belausoy and E. Palevsky, 2009. Interaction of mite Aceria mangifarae and Fusarium mangiferae the causal agent of mango malformation diseases. Phytopathology, 99: 152-159.
Direct Link - Gamliel-Atinsky, E., A. Sztejnberg, M. Maymon, D. Shtienberg and S. Freeman, 2009. Inoculum availability and conidial dispersal patterns of Fusarium mangiferae, the causal agent of mango malformation disease. Phytopathology, 100: 160-166.
CrossRef - Atinksky, E.G., A. Sztejnberg, M. Maymon, H. Vintal, D. Sztejnberg and S. Freeman, 2009. Infection dynamics of fusarium mangiferae causal agent of mango malformation disease. Phytopathology, 99: 775-781.
Direct Link - Azzouz, S., Z.M. Hamdy and I.M. Dahshan, 1978. Studies on malformed inflorescence of mango, the degree of susceptibility among different varieties. Agric. Res. Rev., 57: 17-27.
Direct Link - Bist, L.D. and S. Ram, 1986. Effect of malformation on changes in endogenous gibberellins and cytokinins during floral development of mango. Scientia Hort., 28: 235-241.
CrossRef - Bogale, M., B.D. Wingfield, M.J. Wingfield and E.T. Steenkamp, 2007. Species-specific primers for Fusarium redolens and a PCR-RFLP diagnostic technique for distinguishing among three clades of Fusarium oxysporum. FEMS Microbiol. Lett., 271: 27-32.
PubMedDirect Link - Chakrabarti, D.K. and S. Ghosal, 1985. Effect of Fusarium moniliforme var. subglutinans infection on mangiferin production in the twigs of Mangifera indica. Phytopathol. Z., 113: 47-50.
CrossRef - Chakrabarti, D.K. and S. Ghosal, 1989. The disease cycle of mango malformation induced by Fusarium moniliforme var. subglutinans and the curative effects of mangiferin metal chilates. J. Phytopathol., 125: 238-246.
CrossRef - Chattopadhyay, N.C. and B. Nandi, 1977. Nucleic acid content in malformed mango inflorescence caused by Fusarium moniliforme var. subglutinans. Phytopath. Z., 89: 256-260.
CrossRef - Chattopadhyay, N.C. and B. Nandi, 1978. Changes in to total contents of saccharides, proteins and chlorophyll in malformed mango inflorescence induced by Fusarium moniliforme var. subglutinans. Biol. Plantarum, 20: 468-471.
CrossRef - Chattopadhyay, N.C. and B. Nandi, 1981. Nutrition in Fusarium moniliforme var. subglutinans causing mango malformation. Mycologia, 73: 407-414.
Direct Link - Chib, H.S., P.S. Aandotra and B.R. Gupta, 1984. Survey report on the incidence and extent of mango malformation on mango growing areas of Jammu and Kashmir state. Indian J. Mycol. Plant Pathol., 14: 86-88.
Direct Link - Crowhurst, R.N., B.T. Hawthrone, E.H.A. Rikkerink and M.D. Templeton, 1991. Differentiation of Fusarium solani f. sp. Cucurbitae races1 and 2by Random amplification of polymorphic DNA. Curr. Genet., 20: 391-396.
PubMed - Ghosal, S. and D.K. Chakrabarti, 1988. Differences in phenolic and steroidal constituents between healthy and infected floretes of Mangifera indica. Phytochemistry, 27: 1339-1343.
Direct Link - Ghosal, S., D.K. Chakrabarti, K. Biswas and Y. Kumar, 1979. Toxic substances produced by Fusarium concerning malformation disease of mango. Experimentia, 35: 1633-1634.
CrossRef - Ghosal, S., K. Biswas, D.K. Chakrabarti and K.C. Basuchaudhary, 1977. Control of Fusarium wilt of safflower by mangiferin. Phytopathology, 67: 548-550.
Direct Link - Goodwin, P.H. and S.L. Annis, 1991. Rapid identification of genetic variation and pathotype of Leptosphearia maculans by random ampliphided polymorphic DNA assay. Applied Environ. Microbiol., 57: 2482-2486.
Direct Link - Grajal-Martin, M.J., C.J. Simon and F.J. Muehlbauer, 1993. Use of random amplified polymorphic DNA (RAPD) to characterize race 2 of Fusarium oxysporum f. sp. pisi. Phytopathology, 83: 612-614.
CrossRefDirect Link - Grossman, K., 1996. A role of cyanide derived from ethylene biosynthesis in the development of stress symptoms. Physiol. Plant, 7: 772-775.
CrossRef - Gupta, V.K., A. Pandey, P. Kumar, B.K. Pandey and R.K. Gaur et al., 2010. Genetic characterization of mango anthracnose pathogen Colletotrichum gloeosporioides Penz. by random amplified polymorphic DNA analysis. Afr. J. Biotechnol., 9: 4009-4013.
CrossRefDirect Link - Iqbal, Z., U.M. Rahman, A.A. Dasti, A. Saleem and Y. Zafar, 2006. RAPD of Fusarium islates causing Mango malformation disease in Pakistan. World J. Microb. Biotechnol., 22: 1161-1167.
CrossRef - Jamil, F.F., N. Sarwar, M. Sarwar, J.A. Khan, J. Geistlinger and G. Kahl, 2000. Genetic and pathogenic diversity within Ascochyta rabiei (Pass.) Lab. population in Pakistan causing blight of chikpae (Cicer arientium L.). Physiol. Mol. Plant Pathol., 57: 243-254.
CrossRef - Kelly, A., A.R. Alcala-Jimenez, B.W. Bainbridge, J.B. Heale, E. Perez-Artes and R.M. Jimenez-Diaz, 1994. Use of genetic fingerprinting and random amplified polymorphic DNA to characterize pathotypes of Fusarium oxysporum f.sp. ciceris infecting chickpea. Phytopathology, 84: 1293-1298.
CrossRefDirect Link - Kini, K.R., V. Let and S.B. Mathur, 2002. Genetic variation in Fusarium moniliforme isolated from seeds of different host species from Burkina Faso based on random amplified polymorphic DNA analysis. J. Phytopathol., 150: 209-212.
Direct Link - Kiprop, E.K., J.P. Baudoin, A.W. Mwangombe, P.M. Kimani, G. Mergeal and A. Maquet, 2002. Characterization of Kenyan isolates of Fusarium from Pigeonpea [Cajanus cajan (L.) Millsp.] by cultural characteristics, aggressiveness and AFLP analysis. J. Phytopathol., 150: 517-527.
CrossRef - Kumar, J., U.S. Singh and S.P.S. Beniwal, 1993. Mango malformation: One hundred years of research. Ann. Rev. Phytopathol., 31: 217-232.
Direct Link - Kumar, R. and D.K. Chakrabarti, 1992. Biochemical evidences of physiological specialization of F. moniliforme Sheld, the incidence of malformation disease of M. indica L. Indian J. Expt. Biol., 30: 448-450.
Direct Link - Khush, R.S., E. Becker and M. Wach, 1992. DNA amplification polymorphisms of the cultivated mushroom agaricus bisporus. Applied Environ. Microbiol., 58: 2971-2977.
Direct Link - Kvas, M., E.T. Steenkamp, A.O. Al-Adawi, M.L. Deadman, A.A. Al-Jahwari and W.F.O. Marasas et al., 2008. Fusarium mangifarae associated with mango malformation in Sultanate of Oman. Eur. J. Plant Pathol., 121: 195-199.
CrossRef - Malo, S.E. and R.T. McMillan Jr., 1972. A disease of Mangifera indica L. in Florida similar to mango malformation. Florida State Hort. Soc., 85: 254-268.
Direct Link - Martin-Preveli, P., J. Marchal and V. Furon, 1975. A case of zink deficiency in mango. Fruits, 30: 201-201.
Direct Link - Mishra, K.A. and B.S. Dhillon, 1980. Levels of endogenous gibberellins in the healthy and malformed panicles of mango (Mangifera indica L.) India. Indian J. Hort., 37: 33-34.
Direct Link - Mitter, N., A.C. Srivastava, S. Ahmad, A.K. Sarbhoy and D.K. Agrawal, 2002. Characterization aof gibbereline producing strains of Fusarium moniliforme based on DNA polymorphism. Mycopathologia, 153: 187-193.
Direct Link - Narayanan, E.S. and S. Ghai, 1962. Some new records and a new species of mites associated with malformation of mango trees in India. Proc. Nat. Inst. Sci. India, 29B: 535-546.
Direct Link - Nicholson, R.I.D. and J. van Staden, 1988. Cytokinin and mango flower malformation. I. Tentative identification of the complement in healthy and malformed inflorescence. J. Plant Physiol., 132: 720-724.
Direct Link - Pal, R.N., S.K. Kalra, D.K. Tandon and K.L. Chadha, 1983. Activity of IAA oxidase, catalase and amylase in morphactin-induced malformations of mango inflorescence. Scientia Hortic., 19: 271-277.
CrossRef - Pandey, R.M., M.M. Rao and R.A. Pathak, 1977. Biochemical changes associated with floral malformation in mango. Scientia Hortic., 6: 37-44.
CrossRef - Ploetz, R., Q.I. Zheng, A. Vazquez and M.A. Abdel Sattar, 2002. Current status and impact of mango malformation in Egypt. Int. J. Pest Manage., 48: 279-285.
Direct Link - Raafat, A., S.H. El-Deeb and K.H. Neumann, 1995. Investigations on the endogenous activity of gibbrellins and cytokinins of malformed mango (Mangifera indica L.) in relation to histological disorders. I. Vegitative malformation. Angewandte-Botanic, 69: 3-9.
Direct Link - Raafat, A., S.H. El-Deeb and K.H. Neumann, 1995. Investigation on the endogenous activity of gibberellins and cytokinins of malformed mango (Mangifera indica L.) in relation to histological disorders. II: Floral malformation. Angwandte Botanic, 69: 10-16.
Direct Link - Ram, S. and L.D. Bist, 1984. Occurrence of malformin like substances in malformed panicles and control of floral malformation in mango. Sci. Hort., 24: 331-336.
CrossRef - Riov, J. and S.F. Yang, 1982. Autoinhibition of ethylene production in citrus peel discs. Suppression of 1-aminocyclopropane-1- carboxylic acid synthesis. Plant Physiol., 69: 687-690.
Direct Link - Sharma, I.M. and S.D. Badiyala, 1990. Incidence of mango malformation in different locations of Himanchal Pradesh. Indian J. Mycol. Plant Pathol., 20: 179-181.
Direct Link - Sharma, R.R., H.C. Sharma and A.M. Goswami, 1994. Polyphenol oxidase activity and phenolic content pattern during shoot development of grape (Vitis vinifera L.) in different growing seasons. J. Plant Biochem. Biol., 3: 145-147.
Direct Link - Siefert, F., J. Kwiatokowski, S. Sarkar and K. Grossman, 1995. Changes in endogenous cyanide and 1-aminocyclopropane-1-carboxylic acid levels during the hypersensitive response to tobacco mosaic virus infected tobacco leaves. Plant Growth Regul., 17: 109-113.
CrossRef - Singh, R.N., P.K. Majumder, D.K. Shrama, G.C. Sinha and P.C. Bose, 1974. Effects of deblossoming on productivity of mango. Sci. Hort., 2: 399-403.
CrossRef - Singh, V.K. and R.B.R. Yadav, 1997. Physio-biochemical changes in healthy and clustered fruits of mango cv. Dashehari. Indian J. Plant Physiol., 2: 325-326.
Direct Link - Singh, Z. and B.S. Dhillon, 1989. Nitrogen, protein levels and amino acids composition in developing malformed and healthy panicles and shoots of mango. Acta Hort., 231: 858-865.
Direct Link - Singh, Z. and B.S. Dhillon, 1986. Effect of naphthalene acetic acid, ethrel, dikegulac and hand deblossoming on floral malformation, flowering, yield and fruit quality of mango (Mangifera indica L.). Acta Hort., 175: 307-313.
Direct Link - Singh, Z. and B.S. Dhillon, 1988. Influence of deblossoming on floral malformation of mango (Mangifera indica L.). Trop. Agric., 65: 295-296.
Direct Link - Singh, Z. and B.S. Dhillon, 1989. Relationship of endogenous and exogenous ethylene with floral malformation of mango (Mangifera indicaL.). Acta Hort., 239: 367-370.
Direct Link - Singh, Z. and B.S. Dhillon, 1989. Nitrogen, protein content and amino acids composition of malformed and healthy seedlings of mango. Acta Hort., 231: 878-882.
Direct Link - Singh, Z. and B.S. Dhillon, 1990. In vivo role of Indole-3-acetic acid, gibberellic acid, zeatin, abscisic acid and ethylene in floral malformation of Mangifera indica L. J. Phytopathol., 128: 235-245.
CrossRef - Singh, Z. and B.S. Dhillon, 1990. Comparative developmental morphology of normal and malformed floral organs of mango (Mangifera indicaL.). Tropic. Agric., 67: 143-148.
Direct Link - Singh, Z. and B.S. Dhillon, 1993. Metabolic changes associated with floral malformation of mango (Mangifera indica L.). Tropic. Agric., 70: 68-72.
Direct Link - Singh, Z. and B.S. Dhillon, 1986. Effect of plant growth regulators on floral malformation, flowering productivity and fruit quality of mango (Mangifera indica L.). Acta Hort., 175: 315-319.
Direct Link - Singh, Z. and B.S. Dhillon, 1987. Effect of foliar application of boron on vegetative and panicle growth, sex expression, fruit retention and physiochemical characters of fruits of mango (Mangifera indica L.) cv. Dushehari. Trop. Agric., 64: 305-308.
Direct Link - Singh, Z. and B.S. Dhillon, 1987. Occurrence of malformin like substances in seedlings of mango (Mangifera indica L.). J. Phytopathol., 120: 245-248.
CrossRef - Singh, Z. and B.S. Dhillon, 1989. Hormonal changes associated with vegetative malformation of mango (Mangifera indica L.). J. Phytopathol., 125: 193-197.
CrossRef - Singh, Z. and B.S. Dhillon, 1990. Floral malformation, yield and fruit quality of Mangifera indica L. in relation to ethylene. J. Hort. Sci., 65: 215-220.
Direct Link - Singh, Z. and B.S. Dhillon and C.L. Arora, 1991. Nutrient levels in malformed and healthy tissue of mango (Mangifera indica L.). Plant Sci., 133: 9-15.
CrossRef - Sivramakrisnan, S., S. Kannan and S.D. Singh, 2002. Genetic variability of Fusarium wilt pathogen isolates of chickpea (Cicer arietanum L.) assessed by molecular markers. Mycopathologia, 155: 171-178.
PubMed - Sternlicht, M. and S. Goldenberg, 1976. Mango eriophyid mites in relation to inflorescence. Phytoparasitica, 4: 45-50.
CrossRef - Usha, K., B. Singh, P. Praseetha, N. Deepa, D.K. Agarwal, R. Agarwal and A. Nagaraja, 2009. Antifungal activity of Dhatura stramonium, Calotropis gigantean and Azadirachta indica against Fusarium mangiferae and floral malformation in mango. Eur. J. Plant Pathol., 124: 637-657.
CrossRefDirect Link - Vakalounakis, D.J. and G.A. Fragkiakakis, 1999. Genetic diversity of Fusarium oxysporum isolates from cucumber: Differentiation by pathogenicity, vegetative compatibility, and RAPD fingerprinting. Phytopathology, 89: 161-168.
Direct Link - Van Staden, J., A.D. Bajley and S. Macrae, 1989. Cytokinins and mango flower malformation. III. The metabolism of [H] 3 iso-pentenyladenine and [ 8-14 ] zeatin by Fusarium moniliforme. Physiol. Mol. Plant Pathol., 35: 433-438.
CrossRef - Welsh, J. and M. McClelland, 1990. Fingerprinting genomes using PCR with arbitrary primers. Nucleic Acids Res., 18: 7213-7218.
CrossRefPubMedDirect Link - Williams, J.G.K., A.R. Kubelik, K.J. Livak, J.A. Rafalski and S.V. Tingey, 1990. DNA polymorphisms amplified by arbitrary primers are useful as genetic markers. Nucleic Acids Res., 18: 6531-6535.
CrossRefPubMedDirect Link - Wilson, A., D. Simpson, E. Chandler, P. Jennings and P. Nicolson, 2004. Development of PCR assays for the detection and differentiation of Fusarium sporotrichiodes and Fusarium langsethiae. FEMS Microbiol. Lett., 233: 69-76.
CrossRef - Youssef, S.A., M. Maymon, A. Zveibil, D. Klein-Gueta, A. Sztejnberg, A.A. Shalaby and S. Freeman, 2007. Epidemiological aspects of mango malformation disease caused by Fusarium mangiferae and source of infection in seedlings cultivated in orchards in Egypt. Plant Pathol., 56: 257-263.
Direct Link - Zheng, Q. and R. Ploetz, 2002. Genetic diversity in the mango malformation pathogen and development of a PCR assay. Plant Pathol., 51: 208-216.
Direct Link - Miller, J.M. and E.E. Conn, 1980. Metabolism of hydrogen cyanide by higher plants. Plant Physiol., 65: 1199-1202.
PubMedDirect Link - Kaemmer, D., R. Afza, K. Weisang, G. Kahl and F.J. Novak, 1992. Oligonucleotide and amplification fingerprinting of wild species of wild species and cultivars of bananas (Musa sp.). Biotechnology, 10: 1030-1035.
CrossRef - Jana, T., T.R. Sharma, R.D. Prasad and D.K. Arora, 2003. Molecular characterization of macrophomina phaseolina and Fusarium species by a single primer RAPD technique. Microbiol. Res., 158: 249-257.
PubMedDirect Link - Miller, S.A., 1996. Detecting propagules of plant pathogenic fungi. Adv. Bot. Res., 23: 73-102.
CrossRef