
S.A. Shameem
Division of Environmental Sciences, SKUAST-Kashmir, Srinagar (J&K), Forest Research Institute University Dehradun, Uttarakhand, India
P. Soni
Ecology and Environment Division, Forest Research Institute University, Dehradun, Uttarakhand, India
G.A. Bhat
Department of Environmental Sciences/CORD, University of Kashmir, Srinagar (J&K), India
Asian Journal of Plant Sciences
Year: 2010 | Volume: 9 | Issue: 6 | Page No.: 329-336







ABSTRACT
The present study was conducted to estimate the variation in herbaceous community features in terms of diversity, species richness and distribution pattern in the two different ecosystems i.e., site I (pastureland) and site II (forest) in the lower Dachigam National Park of Kashmir, Himalaya. The pasture site is located outside the National Park and is under grazing were as forest site is located inside the National Park and is protected. The study was done on seasonal basis and the results revealed higher trend for diversity(H') at both sites during summer season (site I = 3.03, site II =2.92). However, dominance index was inversely related to diversity index(H') (site I = 0.06 and site II = 0.07). Richness index showed maximum value in spring (4.12, site II) and summer (3.70, site I). Equability or evenness index obtained higher value in summer (0.94, site I) and winter (0.96, site II). Comparatively on an average site II showed higher trend in Shannon diversity (site I = 2.435, site II = 2.655), richness index (site I = 2.652, site II = 3.297) and equability index (site I = 0.852, site II = 0.915). The frequently occurred dominant species during prominent season based on IVI at site I were Cynodon dactylon, Salvia moorcroftiana and Thymus serphyllum at site II were Arthraxon prinoides, Fragaria nubicola, Poa annua, Poa sp. and Viola indica. The abundance to frequency ratio (A/F) indicated most of the species performed contiguous pattern of distribution. A seasonal picture depicted contagious> random distribution pattern while regular distribution was reported almost negligible. The study concluded that seasons have great influence on species diversity. During spring and summer season an increase in species diversity was observed which declined as autumn and winter approached mainly due to dry environmental conditions, slow growth rate and other climatic factors.
PDF Abstract XML References Citation
Received: March 05, 2010;
Accepted: July 16, 2010;
Published: September 20, 2010
How to cite this article
S.A. Shameem, P. Soni and G.A. Bhat, 2010. Comparative Study of Herbaceous Vegetation in Lower Dachigam National Park, Kashmir Himalaya, India. Asian Journal of Plant Sciences, 9: 329-336.
DOI: 10.3923/ajps.2010.329.336
URL: https://scialert.net/abstract/?doi=ajps.2010.329.336
DOI: 10.3923/ajps.2010.329.336
URL: https://scialert.net/abstract/?doi=ajps.2010.329.336
INTRODUCTION
The destruction of vegetation has been continuing at an alarming pace world over due to a variety of causes (Prance et al., 2000; Pimm et al., 1995). It is estimated that about 8% of the scientifically known plant species on earth are presently on the brink of extinction (Chapin et al., 2000). At global level, concerns about the changes in land use/ cover emerged due to realization that land surface processes influence climate and that change in these processes impact on ecosystem goods and services (Lambin et al., 2003). The impacts that have been of primary concern are the effects of land use change on biological diversity, soil degradation and the ability of biological systems to support human needs. Crop yields have declined, forcing people to cultivate more and more land to meet their needs (Kaihura and Stocking, 2003).
Distinguishing plant communities has been at the heart of vegetation science for centuries, with a traditional focus on the distribution, composition and classification of plant communities (Kashian et al., 2003). Plant communities are defined as an assemblage of functionally similar species populations that occur together in time and space (Magurran, 1988). Plant communities are separated from each other based on indicator species in combination with a distinctive floristic composition. Therefore, any depletion in biodiversity is bound to alter the community attributes (Mishra et al., 2004). Generally, biodiversity measurement typically focuses on the species level and species diversity is one of the most important indices which are used for the evaluation of ecosystems at different scales (Ardakani, 2004).
Biological diversity implies the variety of living organisms which includes diversity within species, between species and of ecosystems and the ecological processes of which they are a part (Gaston and Spicer, 1998). Globally, biodiversity is changing at an unprecedented rate as a complex response to several human-induced changes (Vitousek, 1994; Hooper et al., 2005). These changes in biodiversity cause concern for ethical, economical, ecological and aesthetic reasons, but they also have a strong potential to alter ecosystem services such as the prevention of soil erosion and maintenance of hydrologic cycles and ecosystem goods, like tourism and recreation. Beyond the ecosystem services, biodiversity influences many ecosystems properties such as productivity, decomposition rates, nutrient cycling and resistance and resilience to perturbations (Loreau et al., 2001). Moreover, a high biodiversity is seen as an insurance against a decline in ecosystem services and should therefore be preserved (Yachi and Loreau, 1999).
The structure of plant as well as animal communities in many natural ecosystems is largely influenced by the disturbances, frequently occurring in the system naturally or due to anthropogenic activities (Bennett and Adams, 2004; Eldered and Doak, 2006; Kwit and Platt, 2003). In many of these systems, disturbances change overall community structure (Shaforth et al., 2002) which in turn can ultimately affect community and population dynamics. In other words, the global environmental degradation has been severely occurred and has been introduced as one of the main environmental troubles worldwide. Studying vegetation and various environmental factors (e.g., physiographic, climate, soil, etc.), the community stability and the factors correlation with the vegetation can be reached, which is crucial in terms of forest communities development and rehabilitation (Basiri, 2003). Grazing areas have become less and less productive resulting from over stocking of livestock. Conflicts over the use of land have increased due to increased demand for land by different sectors of the economy. Of particular concern are the conflicts among cultivators, livestock keepers, wildlife conservationists, individual land users and governments due to encroachment of humans into the protected areas (Hoare, 1999; Campbell et al., 2003; Wells and Brandon, 1992). Increased plant species diversity has been linked to improvements in ecosystem function, including increased primary (plant) productivity, greater stability in response to disturbance, improved nutrient cycling and greater resistance to weed invasion (Hector et al., 1999; Spehn et al., 2005). These improvements in function could be of great benefit to agriculture (Tilman et al., 1999; Minns et al., 2001). Preliminary research suggests that manipulating plant diversity can improve primary production in grazed systems (Sanderson et al., 2005) and can reduce weed pressure (Tracy and Sanderson, 2004). The role of plant species diversity in pastures, mainly from a plant and soil viewpoint, was reviewed by Sanderson et al. (2004). Due to increasing human population, the biotic pressure on native forest is inevitable and their unprecedented exploitation is continuously increasing day by day thus reducing the available resource for future use to humans and other life forms who dependent directly or indirectly on such ecosystems.
The herbaceous layer composition is changing continuously in space and time due to multitude of factors such as grazing, fire and rainfall which differs in intensity and duration. Kashmir Himalaya, due to its rich repository of vegetation has attracted naturalists and botanists for more than two centuries (Dar et al., 2001). Numerous studies dealing with diverse aspects of vegetation from different areas of the region have been carried out from time to time (Stewart, 1982; Dar et al., 2001). The general vegetation of Dachigam has been dealt in detail by Singh and Kachroo (1976). They have recognized a number of vegetational types based on habitat, form and density of dominant species, though the vegetation patterns are controlled by such factors as habitat, slope, exposure to sunlight and altitude, besides biotic factors. Anthropogenic disturbances and livestock grazing can adversely affect the composition of herbaceous vegetation in different ecosystems it therefore became imperative to conserve the herbaceous vegetation of the two selected sites.
MATERIALS AND METHODS
Study site: Dachigam National park is located between 34°. 04' -34°. 11' N -latitude and 74°. 54'-75°. 09' E-longitude is about 20 km away from Srinagar, Kashmir with an undulating mountain valley topographic system. The entire area of the park is distinguishable into two sectors upper and lower Dachigam which is spread over an area of 141 km2. The present study was confined to the lower Dachigam conducted during the year 2002-2006 at two different ecosystems viz., site-I (pastureland located outside the official boundary of the Park) and site-II (forest located inside the official boundary of the Park).
Methodology: Vegetation survey of plant communities was conducted in the selected sites during four prominent seasons Spring (March-May), Summer (June-August), Autumn (September-November) and Winter (December-February) using permanent 1mx1m quadrats. Twenty five quadrats were laid out randomly using quadrat method at the two selected sites to workout various community features (Sharma et al., 1983; Rajvanshi et al., 1987). The vegetation data recorded was quantitatively analysed for density, frequency and abundance following Curtis and McIntosh (1950). The relative values of these indices were determined as per Phillips (1959). These values were summed up to get important value index (IVI) of individual species (Curtis, 1959). Diversity index (H') of each sample stand was calculated using the method prescribed by Shannon and Wiener (1963) and Concentration of Dominance (CD) was calculated following the formula given by Simpson (1949). Evenness or equability index was calculated according to Pielou (1966) whereas richness index (D) was calculated according to Margalef (1958). The ratio of Abundance to Frequency (A/F) for different species was determined for eliciting the distribution patterns. This ratio has indicated regular (<0.025), random (0.025-0.05) and contagious (>0.05) distribution (Curtis and Cotton, 1956).
RESULTS
During the study period total number of herbaceous species recorded in site I ranged from 8 in winter to 28 species in summer whereas site II showed a variation of 14 species in winter to 23 each in spring and summer seasons respectively. Thus total number of species encountered at two different sites ranged from 48 in site I (pastureland) to 42 in site II (forest). The seasonal break-up of species recorded during different seasons at two sites are presented in the Fig. 1. Out of the total plant species encountered at both sites (site I = 48 and site II = 42) during four seasons, eighteen highly dominant species based on their IVI are depicted in Fig. 2. Dominant species at site I (pastureland) during different seasons were Salvia moorcroftiana (140.97), Plantago lanceolata (57.56), Thymus serphyllum (41.59), Bothriocloa pertusa (39.51) and Cynodon dactylon (37.29) whereas at site II(forest) Poa steralis (72.96), Poa sp. (56.80),
Viola indica (45.06) Oxalis corniculata (38.72), Fragaria nubicola (30.75) and Viola odorata (30.40) showed higher dominance. Different diversity indices used to estimate the herbaceous structure of site I are presented in the Fig. 3. The results revealed that species diversity (H') recorded at this site showed a range of 1.94-autumn to 3.03-summer seasons. Dominance showed opposite trend to that of Shannon diversity (0.25-autumn to 0.06-summer). Evenness or Equability index showed an increasing trend from spring onwards reaching maximum (0.94) in summer and thereafter decreasing with a minimum value (0.73) during autumn season. Spring and summer season showed highest trend in species richness (3.04, 3.70) followed by a decreased trend in autumn (2.07) reached lowest at winter season (1.87). Different diversity indices recorded at site II are presented in Fig. 4. The obtained observation for diversity index (H') recorded for this site showed spring and summer season at higher side (2.81, 2.92) followed by autumn and winter (2.36, 2.53). Dominance showed reverse trend to that of Shannon diversity (0.07, summer and 0.13, autumn). Equability or evenness index showed similar trend in their approach to that of Shannon-Weiner diversity at site II with spring and summer season at higher side (0.90, 0.93) followed by autumn and winter falling at lower side (0.87, 0.96).
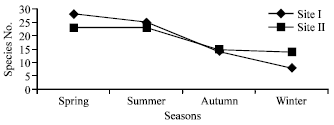 | |
| Fig. 1: | Species recorded during different seasons at two sites |
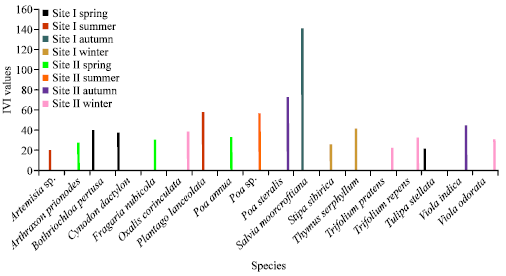 | |
| Fig. 2: | Eighteen highly dominant herbaceous plant species recorded at two sites during different seasons |
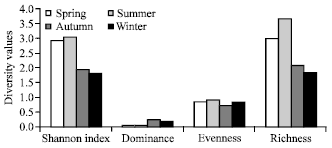 | |
| Fig. 3: | Different diversity estimates of the herbaceous species in site I during different seasons |
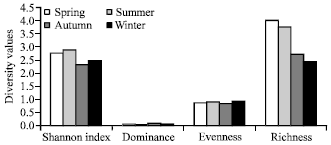 | |
| Fig. 4: | Different diversity estimates of the herbaceous species in site II during different seasons |
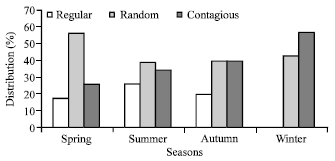 | |
| Fig. 5: | Distribution pattern (%) of herbaceous species at site I during different season |
Species richness at this site was observed maximum during spring season (4.12) and thereafter a decreasing trend was seen reaching minimum richness in winter season (2.48). However, average diversity, evenness and richness index showed slightly higher trend at site II (forest). Common species encountered between two sites during different seasons include Oxalis corniculata (spring to autumn), Stipa sibirica (spring and summer), Trifolium pratens (spring and summer), Trifolium repens (spring and summer), Tulipa stellata (spring), Lychnis coronaria (autumn) and Verbasium Thapsus (spring) season.
Abundance and frequency ratio (A/F) showed a contagious distribution pattern followed by random at both sites during different seasons. At site I about 26.09% species during spring and 57.15% species in winter season showed contagious distribution followed by 39.14% species in summer to 56.53% species during winter showing random distribution (Fig. 5).
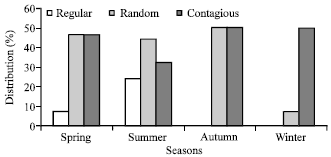 | |
| Fig. 6: | Distribution pattern (%) of herbaceous species at site II during different seasons |
At site II 32% to 50% species showed contagious distribution during summer to autumn-winter season followed by 7.15% (winter) to 50% species (autumn) season showed random distribution (Fig. 6). However, regular distribution at both sites was reported as negligible.
DISCUSSION
Diversity is a combination of two factors-the number of species present, referred to as species richness and the distribution of individuals among species, referred to as species evenness or equability. Species diversity therefore, refers to the variation that exists among the different life forms. The general structure of species at both sites indicated increasing trend in their number mostly during spring and summer season (site I, Spring = 28; Summer = 25), (site II, Spring = 23; Summer = 23). The reason for their maximum occurrence during these two season could be due to availability of moisture provided most oftenly by rains and through other environmental factors. Similar pattern of observations mirrored to present study were also mentioned by Sharma and Upadhyaya (2002). The species diversity in the present study ranged from (1.84-3.03) at pasture site and (2.36-2.92) in forest site. An increasing trend in species diversity was observed from spring onwards which declined with the commencement of autumn and winter at both sites. Comparatively, results of Shannon diversity(H') fall within the range of the study carried out by Kiss et al. (2004) and Yadav and Gupta (2007). However, pasture site showed highest species diversity during summer season which might be due to moderate disturbance by grazing and invasion of new species. Decocq et al. (2004) also reported species diversity highest in intermediate disturbance ecosystem than in undisturbed forests. Many other studies mentioned similar observations pertaining to the present study emphasizing that moderate level of grazing promoted species diversity (Rikhari et al. (1993); Singh et al. (2003). However, Sher et al. (2005) suggested that protection from grazing and over-harvesting for few years could increase plant diversity in pasturelands. Hegland et al. (2001) and Sher et al. (2005) reported that protection might help in the revival of original vegetation and recovery of natural habitats where degradation resulted due to heavy grazing and other biotic pressure. The lower diversity during autumn and winter at both sites could be due to lower rate of evolution and diversification of communities (Fischer, 1960; Simpson, 1964) and severity in environment (Connel and Oris, 1964). Concentration of dominance ranged from (0.07-0.13) in forest site and (0.06-0.25) in pasture site hence inversely related to diversity(H'). The lower value of dominance at forest site shows that dominance of herb layer plants is shared by many species. The Pielou’s indices at both sites were around (0.915-0.0825) on average, indicating low dominance and more or less regular distribution of plant species at both sites. Index of richness was recorded minimum during autumn and winter seasons which could be due to dry environmental conditions and also due to slow growth rate, to a maximum during spring and summer at both sites only due to favourable climatic conditions. High IVI value of a species indicated its dominance and ecological success, its good power of regeneration and greater ecological amplitude. It does vary with the season. The reason that certain species grow together in a particular environment is usually because they have similar requirement for existence in terms of environmental factors such as light, temperature, water and soil nutrients and drainage etc. They may also share the ability to tolerate the activities of animals and humans such as grazing, burning, cutting or trampling (Wood et al., 1994). In accordance to this at site I (pastureland) Salvia moorcroftiana and Thymus serphyllum showed maximum IVI value during autumn and winter season indicating its dominance due to environmental suitability and ability of the species against grazing during these two seasons. Moreover, their dominance at the site could possibly be due to availability of optimum conditions for their growth. Similar observations were also reported by Kukshal et al. (2009) based on seasonal changes among species in their IVI that makes them dominant different during seasons. The growing dominance of non-palatable species and others in the pasture site is probably an indication of adaption against herbivory and adverse climatic conditions. Bhandari et al., (1999) while working in pasturelands of Garhwal Himalaya reported favourable results in concurrent to the present study. Moreover higher value of IVI by any individual species indicates that all the available resources are being utilized by that species and left over are being trapped by another species as the competitors and the associates. This could be the reason why IVI was found to be always highest among few species during autumn than rest of the three seasons could be most oftenly as number of species went on decreasing as the autumn approaches and thus the relative IVI gets shared among few individual species thereby resulted higher IVI among them at both sites. In this context it can be correlated that the species (Arthraxon prinoides, Fragaria nubicola, Poa annua, Poa sp. and Viola indica) presented maximum IVI values at site II during most of the season. Furthermore the distribution of niche space or availability of the resource was equally distributed among all species during most of the seasons (both sites) while in rest of the season it showed a reverse trend that is why only few species showed high dominance during that period thus occupied more niche space than rest of the species. It is also to mention that the dominance of certain species at both sites in a particular period is possible mainly as the other co-dominant species do not reach maturity to complete their life cycle. Similar kind of view parallel to the present study were also reported by Vesk and Westoby (2000) and Sher et al. (2005).
Abundance and frequency ratio (A/F) ratio was used to assess the distribution pattern of species. It reveals that most of the species were contagiously distributed whereas as regular distribution was reported at lower side during different seasons. The results reported by Shadangi and Nath (2005) showed maximum species in contagious distribution. Ilorkar and Khatri (2003) investigated herb layer species in contagious distribution followed by random. Contagious distribution pattern in forest ecosystem was also reported by Kumar et al. (2004) and Chen et al. (2008) hence supports the results obtained in present study. Dominance of contagious distribution may be due to the fact the majority of species reproduce vegetatively in addition to their sexuality. Odum (1971) described that in natural conditions contagious distribution is most common type of distribution and is performed due to small but significant variation in environmental conditions, while random distribution is found only in very uniform environment. Contagious distribution in natural vegetation has been reported by Greigh-Smith (1957), Kershaw (1973), Singh and Yadava (1974) confirms the results of the present study within their optimum range. However, observations indicated that contagious distribution in vegetation (as recorded for both sites) was due to multitude factors and the vegetative reproduction may not be the only reason (Kershaw, 1973; Saxena and Singh, 1982).
CONCLUSION
From the present investigation, it can be inferred that diversity of ground flora is closely related with variations in various environmental variables. Species diversity increases during spring season and declines thereafter. This character is attributed to the fact that during spring/ summer season new species go on sprouting depending upon the root/ seed stock in the soil and thereby adding to species in total resulted more diversity. During autumn and winter the rate of sprouting of root/ seed stock is diminished and species number declined owing to adverse climatic conditions which ultimately reduces species diversity. It is also to be concluded that the biotic interfere at both sites need to be monitored regularly on seasonal basis which will certainly enhance species diversity of herbaceous vegetation at two selected sites.
ACKNOWLEDGMENTS
The authors are grateful to the Director Forest Research Institute Dehradun, India for allowing to conduct the Research work in the Dachigam National Park. Director CORD/ P. G. Department of Environmental Science University of Kashmir is highly acknowledged for providing laboratory facility. Thanks are also due to Chief Wildlife Warden, J and K Government for allowing to use the National Park for research purpose.
REFERENCES
- Bennett, L.T. and M.A. Adams, 2004. Assessment of ecological effects due to forest harvesting: Approaches and statistical issues. J. Applied Ecol., 41: 585-598.
Direct Link - Bhandari, B.S., D.C. Nautiyal and R.D. Gaur, 1999. Structural attributes and productivity potential of an alpine pastures of Garhwal Himalaya. J. Indian Bot. Soc., 78: 321-329.
Direct Link - Chapin, F.S., E.S. Zavaleta, V.T. Eviner, R.L. Naylor and P.M. Vitousek et al., 2000. Consequences of changing biodiversity. Nature, 405: 234-242.
CrossRefDirect Link - Connel, J.H. and E. Oris, 1964. The ecological regulation of species diversity. Am. Nat., 48: 399-414.
Direct Link - Gaston, K.J. and V.I. Spicer, 1998. Biodiversity: An Introduction. 2nd Edn., Blackwell Science, Oxford, UK., ISBN-13: 978-1405118576.
Direct Link - Hoare, R.E., 1999. Determinants of human-elephant conflict in a landuse mosaic. J. Applied Ecol., 36: 689-700.
Direct Link - Hooper, D.U., F.S. Chapin III, J.J. Ewel, A. Hector and P. Inchausti et al., 2005. Effects of biodiversity on ecosystem functioning: A consensus of current knowledge. Ecol. Monogr., 75: 3-35.
CrossRefDirect Link - Ilorkar, V.M. and P.K. Khatri, 2003. Phytosociological study of Navegaon National Park, (Maharashtra). Indian For., 129: 377-387.
CrossRefDirect Link - Kashian, D.M., B.V. Barnes and W.S. Walker, 2003. Ecological species groups of landform level ecosystems dominated by jack pine in northern Lower Michigan, USA. Plant Ecol., 166: 75-91.
CrossRef - Kukshal, S., B.P. Nautiyal, A. Anthwal, A. Sharma and A.B. Bhatt, 2009. Phytosociological investigation and life form pattern of grazinglands under pine canopy in temperate zone, Northwest Himalaya, India. Res. J. Bot., 4: 55-69.
CrossRefDirect Link - Kwit, C. and W.J. Platt, 2003. Disturbance history influences regeneration of non-pioneer under storey trees. Ecology, 84: 2575-2578.
Direct Link - Lambin, E.F., H.J. Geist and E. Lepers, 2003. Dynamics of land use and land cover change in tropical regions. Ann. Rev. Environ. Res., 28: 205-241.
Direct Link - Loreau, M., S. Naeem, P. Inchausti, J. Bengtsson and J.P. Grime et al., 2001. Biodiversity and ecosystem functioning: Current knowledge and future challenges. Science, 294: 804-808.
CrossRefDirect Link - Mishra, B.P., O.P. Tripathi, R.S. Tripathi and H.N. Pandey, 2004. Effects of anthropogenic disturbance on plant diversity and community structure of a sacred grove in Meghalaya, Northeast India. Biodiversity Conserv., 13: 421-436.
CrossRef - Pimm, S.L., G.J. Russell, J.L. Gittleman and T.M. Brooks, 1995. The future of biodiversity. Science, 269: 347-350.
CrossRefPubMedDirect Link - Prance, G.T., H. Beentje, J. Dransfield and R. Johns, 2000. The tropical flora remains undercollected. Ann. Missiri Bot. Gard., 87: 67-71.
Direct Link - Rikhari, H.C., G.C.S. Negi, J. Ram and S.P. Singh, 1993. Human-induced secondary succession in alpine meadow of Central Himalaya, India. Arctic Alpine Res., 25: 8-14.
Direct Link - Saxena, A.K. and J.S. Singh, 1982. A phytosociological analysis of woody species in forest communities of a part of Kumaon Himalaya. Vegetatio., 50: 3-22.
CrossRefDirect Link - Sharma, K.P. and B.P. Upadhyaya, 2002. Phytosociology, primary production and nutrient retention in herbaceous vegetation of the forestry arboretum on the Aravalli hills at Jaipur. Trop. Ecol., 42: 325-335.
Direct Link - Singh, J.B. and P. Kachroo, 1976. Forest Flora of Srinagar and Plants of Neighbourhood. 1st Edn., Bishen Singh Mahendra Pal Singh, Dehradun, India, ISBN: 81-7622-131-7.
Direct Link - Singh, J.S. and P.S. Yadava, 1974. Seasonal variation in composition, plant biomass and net primary productivity of a tropical grassland at Kurukshetra. India Ecol. Monogr., 44: 351-375.
CrossRef - Wood, J., A.B. Low, J.S. Donaldson and A.G. Rebelo, 1994. Threats to Plant Species Diversity Through Urbanization and Habitat Fragmentation in the Cape Metropolitan Area, South Africa. In: Strelitzia 1 Botanical Diversity in Southern Africa, Huntley, B.J. (Ed.). National Botanical Institute, UK., pp: 259-274.
- Yadav, A.S. and S.K. Gupta, 2007. Effect of micro-environment and human disturbance on the diversity of herbaceous species in Sariska Tiger project. Trop. Ecol., 48: 125-128.
Direct Link - Yachi, S. and M. Loreau, 1999. Biodiversity and ecosystem productivity in a fluctuating environment: The insurance hypothesis. Proc. Nat. Acad. Sci. USA., 96: 1463-1468.
Direct Link - Decocq, G., M. Aubert, F. Dupont, D. Alard, R. Saguez and A. Wattez-Franger et al., 2004. Plant diversity in a managed temperate deciduous forest: understorey response to two silvicultural systems. J. Applied Ecol., 41: 1065-1079.
Direct Link - Hector, A., B. Schmid, C. Beierkuhnlein, M.C. Caldeira and M. Diemer et al., 1999. Plant diversity and productivity experiments in European grasslands. Science, 286: 1123-1127.
CrossRefPubMedDirect Link - Hegland, S.J., M. van Leeuwen and J.G.B. Oostermeijer, 2001. Population structure of Salvia pratensis in relation to vegetation and management of Dutch dry floodplain grassland. J. Applied Ecol., 38: 1277-1289.
Direct Link - Minns, A., J. Finn, A. Hector, M.C. Caldeira and J. Joshi et al., 2001. The functioning of European grassland ecosystems: Potential benefits of biodiversity to agriculture. Outlook Agric., 30: 179-185.
Direct Link - Spehn, E.M., A. Hector, J. Joshi, M. Scherer-Lorenzen and B. Schmid et al., 2005. Ecosystem effects of biodiversity manipulations in European grasslands. Ecol. Monogr., 75: 37-63.
CrossRefDirect Link - Vitousek, P.M., 1994. Beyond global warming: Ecology and global change. Ecology, 75: 1861-1876.
Direct Link - Tracy, B.F. and M.A. Sanderson, 2004. Forage productivity, species evenness and weed invasion in pasture communities. Agric. Ecosyst. Environ., 102: 175-183.
CrossRef