
Bo Guan
Laboratory of Coastal Wetland Ecology, Yantai Institute of Coastal Zone Research,Chinese Academy of Sciences, Yantai, 264000, The People�s Republic of China
Junbao Yu
Laboratory of Coastal Wetland Ecology, Yantai Institute of Coastal Zone Research,Chinese Academy of Sciences, Yantai, 264000, The People�s Republic of China
Zhaohua Lu
Laboratory of Coastal Wetland Ecology, Yantai Institute of Coastal Zone Research,Chinese Academy of Sciences, Yantai, 264000, The People�s Republic of China
Wisdom Japhet
Department of Biological Sciences, Ahmadu Bello University, Zaria Kaduna State, Nigeria
Xiaobing Chen
Laboratory of Coastal Wetland Ecology, Yantai Institute of Coastal Zone Research,Chinese Academy of Sciences, Yantai, 264000, The People�s Republic of China
Wenjun Xie
Research Center for Eco-Environmental Sciences Yellow River Delta,Binzhou University, Binzhou, 256603, The People`s Republic of China
Asian Journal of Plant Sciences
Year: 2010 | Volume: 9 | Issue: 4 | Page No.: 194-199







ABSTRACT
The main aim of this study is to detect the effects of NaCl, NaHCO3 and sea salt on seed germination, seedling growth and seedling cation contents of two Suaeda species. Three germination experiments of S. corniculata and S. salsa seeds were conducted in growth chambers. The seeds were placed at three types of salt at concentrations: 0, 25, 50, 75, 100% seawater; 0, 100, 200, 300, 400, 500 mM NaCl and 0, 100, 200, 300, 400, 500 mM NaHCO3, separately. The two species varied in their salt tolerance for germination rates and percentages and showed higher germination percentage at higher salt stress (500 mM NaCl, NaHCO3 and 100% seawater). Some un-germinated seeds were recovered after being transferred to distilled water. The Na+ content in seedlings increased with the increase in stress intensity. While K+ content and K+/Na+ ratio decreased under NaCl and NaHCO3 stress. K+ content increased in seawater treatment while reaching higher salt concentrations, due to the extra K+ in seawater, but there was no significant difference among the treatments with varied seawater concentrations (p<0.001). These results suggested that S. corniculata and S. salsa could be used as pioneer plants for ecological recovery and exploitation of saline and sodic soils.
PDF Abstract XML References Citation
Received: March 15, 2010;
Accepted: May 28, 2010;
Published: June 19, 2010
How to cite this article
Bo Guan, Junbao Yu, Zhaohua Lu, Wisdom Japhet, Xiaobing Chen and Wenjun Xie, 2010. Salt Tolerance in Two Suaeda Species: Seed Germination and Physiological Responses. Asian Journal of Plant Sciences, 9: 194-199.
DOI: 10.3923/ajps.2010.194.199
URL: https://scialert.net/abstract/?doi=ajps.2010.194.199
DOI: 10.3923/ajps.2010.194.199
URL: https://scialert.net/abstract/?doi=ajps.2010.194.199
INTRODUCTION
Over 800 million hectares of land are affected by salinity throughout the world (Munns, 2005). Saline soils formed when evaporation greatly exceeds precipitation for at least part of the year and where salts are present in moderate to high amounts in the parent material of the soil or with a saline water table at shallow depth. Apart from their high salt content saline soils show a considerable diversity in their hydrological, physical and chemical properties. Saline soils may be calcium dominated, sodium dominated or magnesium dominated with a subsequent tendency towards structural degradation (depending on the presence or absence of calcium) (http://www.fao.org/AG/agL/agll/prosoil/saline.htm). Under extreme climatic conditions (low rainfall, high evaporation) salts which are present in soil solution can precipitate at the surface in various forms such as white efflorescence, salt crusts and so on (http://www.fao.org/AG/agL/agll/prosoil/ saline.htm).
Salt accumulation may limit plant growth to salt tolerant plants (halophytes) only. Tolerance also varies with stages of their life cycle, which could be expressed as: (1) the ability to tolerate high salinity without loosing viability while stored in the soil (seed bank), (2) the ability to germinate at high salinities and (3) the ability to complete its life cycle at high salinities (Khan and Gul, 2002). Ravindran et al. (2007) have demonstrated that soil salinity levels, as measured by Electrical Conductivity (EC), can be reduced by the cultivation of halophytes which are able to accumulate salts in their plant tissues on soils affected by salinity. Among them, Suaeda corniculata is a halophyte which often occurs in sodic soils in plains, such as Songnen Plain, where sodium chloride (NaCl) and sodium bicarbonate (NaHCO3) are the most abundant salts (Wang et al., 2007). Leaves of S. corniculata from saline-sodic inland are green during the whole growing period (http://www.efloras.org/florataxon.aspx?flora_id=2 and taxon_id=200006943). S. salsa is a typical halophyte that naturally occurs in highly saline soils in coastal saline wetland (Wang et al., 2004). S. salsa in costal wetland is usually submerged completely in seawater and the salt concentrations in the soil are always high during seed germination and at the seedling stage (Song et al., 2008). Leaves and stems of S. salsa in this area are red-violet during the whole growth period, Fresh branches of S. salsa are very valuable as a vegetable and the seeds can produce edible oil (Zhao et al., 2002). These two species (Chenopodiaceae) are common in saline soils, acquisition and maintenance of high water content may be the strategies these species use to accumulate Na+ which may substitute for an osmoticum or regulate internal ion concentrations under saline conditions.
Seed germination is the initial and most crucial stage in the life cycle of plants (Grime and Campbell, 1991). Different abiotic factors such as temperature, soil salinity, photoperiod and soil moisture affect germination of halophytes (Noe and Zedler, 2000; Khan, 2003; Guan et al., 2009). However, the effect of soil salinity seems to dominate over all other factors in saline areas (Keiffer and Ungar, 1997; Heidari, 2009). The ability for seeds to recover in non-saline solution following transfer from saline solution also plays an important role in saline areas, which has been reported for several halophytes (Ungar, 1978; Khan and Ungar, 1984; Hanslin and Eggen, 2005).
High Na+ tissue content has often been considered as the major factor responsible for salt toxicity in plants. It is conventionally assumed that an ability to exclude Na+ correlates with plant salt tolerance (Munns and James, 2003; Garthwaite et al., 2005). However, the importance of maintaining an optimal K+/Na+ ratio for plant salt tolerance is hardly surprising and is well discussed in literatures (Cuin et al., 2003; Chen et al., 2007). It is also obvious that such an optimal ratio can be maintained by either restricting Na+ accumulation in plant tissues or by preventing K+ loss from the cells (Chen et al., 2007).
The aim of this study were (1) to investigate the seed germination of two Suaeda species treated with NaCl, NaHCO3 and the sea salts, (2) to discuss the physiological responses of Suaeda species to different salts at germination stage.
MATERIALS AND METHODS
Seed collection site: Seeds of S. corniculata were obtained from Songnen Plain (44°3'N, 124°3'E) in 2008, where the soils are highly saline. Seeds of S. salsa were collected from the coastal wetland in Yellow River Delta (37°35'-38°12'N, 118°33'-119°20'E) in the autumn of 2008. The seeds were stored in paper bags at room temperature (20±3°C) until the experiment in January 2009.
Germination experiments: This study was carried out at Laboratory of Coastal Wetland Ecology, Yantai Institute of Coastal Zone Research, Chinese Academy of Sciences, China from January till May in 2009. Seeds were surface-sterilized in 1% sodium hypochlorite solution for 15 min, rinsed in distilled water and dried before the experiment. The seeds were germinated in petri dishes (10 cm diameter) containing two layers of filter paper with 12 mL of test solution. Germination tests were carried out in growth chambers (BSG-800, Shanghai, China) with a 12 h photoperiod (Sylvania cool white fluorescent lamps, 200 μmol m-2 sec-1, 400-700 nm, 25/15°C). To examine the effects of different salts, we conducted six salinity concentrations (0, 100, 200, 300, 400, 500 mM NaCl), six alkalinity concentrations (0, 100, 200, 300, 400, 500 mM NaHCO3) and five sea water concentrations (0, 25, 50, 75 and 100% seawater). A completely randomized design was used in the germination tests. Four replicates of 50 seeds each were used for each treatment. The seeds were considered to have germinated after radicle emergence. Germination was recorded daily for 10 days (Mokhberdoran et al., 2009).
Methods of germination expression: The rate of germination was estimated using a modified Timson’s index of germination velocity = ΣG/t, where G is the percentage of seed germination at one-day intervals and t is the total germination period (Khan and Ungar, 1984). The maximum value possible for our data using this index was 100 (i.e., 1000/10). The greater the value, the more rapid is the germination. All seeds from the previous germination tests that did not germinate after 10 days in different salt treatments, were placed in new Petri dishes with filter paper moistened with distilled water and incubated under the same conditions for an additional 10 days to study the recovery of germination. The recovery percentage was determined by the following formula:
where, a is the total number of seeds germinated after being transferred to distilled water, b is the total number of seeds germinated in saline solution and c is the total number of seeds used (modified from Khan and Gulzar, 2003).
Measurement of physiological indices: All seedlings were harvested after 10 days of salt treatment. The seedlings were first washed with distilled water three times and then dried at 60°C to constant weight. Dry samples of plant material (50 mg) were treated with 10 mL deionized water at 100°C for 1 h and the extract was taken to determine free Na+ and K+ concentrations. Inductively coupled plasma atomic emission spectrometry (ICP-AES) were used for the determination of sodium (Na), potassium (K) concentrations. K+/Na+ ratio was then calculated.
Data analysis: Germination data were arcsine transformed before the Analysis of Variance (ANOVA). The data were analysed using SPSS 11.5 (SPSS Inc., Chicago, IL, USA). Experimental data were subjected to one way analysis of variance and the means were separated by the Least Significant Difference (LSD). For the germination data, significance was tested at the 1% level, for the Na and K concentrations, significance was tested at the 0.1% level.
RESULTS
Final germination: The final germination of S. corniculata and S. salsa were significantly affected by NaCl, NaHCO3, or seawater (Table 1) (p<0.01). They both remained unaffected up to 200 mM NaCl and NaHCO3 treatments. In non-saline control treatment, the germination percentages were 80.5 and 91% for S. corniculata and S. salsa, respectively. At the highest NaCl concentration, the percentage of germination was no less than 40%. Moreover, the germination percentage of both S. corniculata and S. salsa were higher than 60% in all NaHCO3 treatments. Only 38% seeds of S. salsa germinated in comparison to 57.5% germination of S. corniculata in 100% seawater treatment (Table 1).
Germination rate: The highest germination rate was observed in non-saline controls and it decreased with the increase of salinity. S. salsa seeds showed higher germination ability than S. corniculata seeds in all treatments except for those treated with 50, 75 and 100% seawater (Table 1).
For the two species, the rate of germination were affected significantly by NaCl treatment and all remained unaffected up to 200 mM NaHCO3 treatments (Table 1) (p<0.01).
Germination recovery: After 10 days of salinity treatment, seeds were transferred to distilled water to determine the recovery of germination. The results presented the un-germinated seeds from all salt treatments recovered. The germination recovery ranged from 5.63% in 500 mM NaHCO3 (but note the original high germination of 64.5%) to 30.43% in 400 mM NaCl for S. corniculata and from 16.22% in 200 mM NaHCO3 to 43.55% in 100% seawater for S. salsa (Table 1).
Physiological indices: Na+ concentrations of the two species increased with increasing NaCl (p<0.001). The overall increasing trends were similar for S. corniculata and S. salsa, except in the treatment of 300 mM NaCl, where Na+ concentrations in S. salsa seedlings were higher than in S. corniculata seedlings (Fig. 1a-f). In NaHCO3 solutions, Na+ concentrations of S. corniculata seedlings increased with increasing NaHCO3 (p<0.001).
| Table 1: | Germination percentage, rate of germination (Mean±SE, n = 4) at different levels of different salt solutions using modified Timson’s index (Khan and Ungar, 1984) and germination recovery using recovery formula (modified from Khan and Gulzar, 2003) |
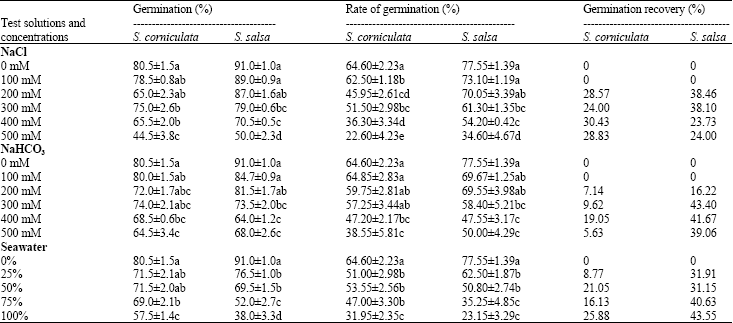 | |
| Different letters indicate significant differences from different concentrations in same salt treatment (p<0.01) | |
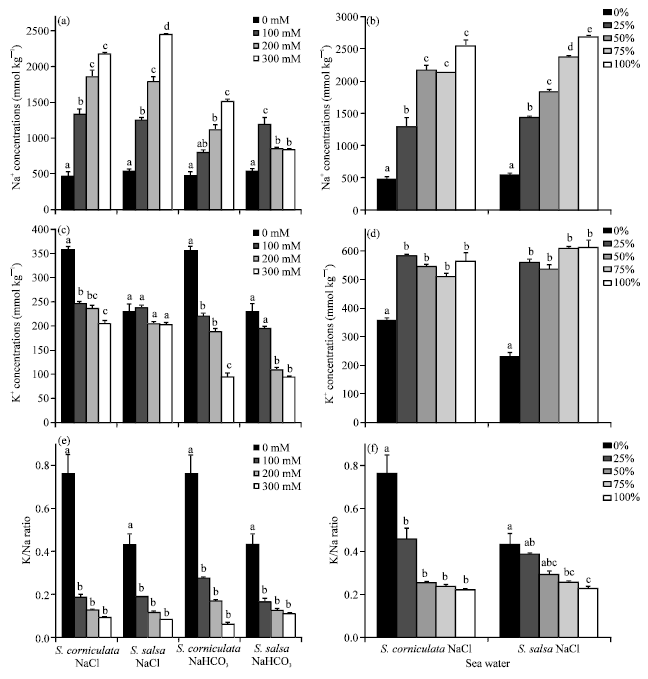 | |
| Fig. 1: | Effects of NaCl, NaHCO3 and seawater on concentrations of Na+, K+ (mmol kg-1 dry mass) and K+/Na+ ratio in the seedlings of S. corniculata and S. salsa. Bars represent±SE (n = 3). Different letters indicate significant differences from each other (p<0.001) |
However, in S. salsa seedlings the concentrations of Na+ decreased with the salinity increased up to 200 mM (Fig. 1). In seawater treatments, Na+ concentrations of S. salsa seedlings increased significantly from 0 to 100% seawater (p<0.001), while they were not significantly different in S. corniculata seedlings treated with 50, 75 and 100% seawater (Fig. 1).
K+ concentrations of S. corniculata seedlings decreased significantly with increasing salinity in both NaCl (p<0.001 and NaHCO3 (p<0.001) treatments (Fig. 1). For S. salsa seedlings, decrease in K+ concentrations was not significant when treated with NaCl (p<0.001). In seawater solutions, K+ concentrations were significantly higher than control (0% seawater), but there was no significant difference among the treatment with different seawater concentrations, which was the case for both S. corniculata and S. salsa seedlings (Fig. 1).
DISCUSSION
The effects of 3 types of salts on seed germination: S. corniculata and S. salsa are typical halophytes which naturally occur in inland saline-sodic plains and coastal saline wetland respectively. For both species, increase in salinity caused a decrease in germination percentage and delayed the germination, but they showed high salt tolerance to NaCl, NaHCO3 and seawater, as quantified by the percentage of germination (Table 1). At 500 mM NaCl, the germination percentage of S. corniculata reached 44.5% and S. salsa reached 50%, this indicated that the two species can establish in most saline environments. This observation was supported by other studies on Arthrocnemum indicum (Khan and Gul, 1998), Salicornia rubra (Khan et al., 2000) and Suaeda japonica (Yokoishi and Tanimoto, 1994). These reports indicate that S. corniculata and S. salsa seeds could be classified as one of the most salt tolerant species during germination. The pH range of NaHCO3 in this study was 8.4 - 9 and the germination percentages of the two species were still high: S. corniculata 64.5% and S. salsa 68% at 500 mM NaHCO3 (Table 1). In sodic soils such as Songnen plain, sodium bicarbonate (NaHCO3) is second most abundant salt next only to sodium chloride (NaCl), high germination percentage makes it promising to use these plants to rebuild the sodic land under serious threat of degradation. Furthermore, seeds of S. corniculata and S. salsa are highly salt tolerant and can germinate at 100% seawater salinity. This may follow the same rule as combined salt effect because in seawater a lot of ions were included (Duan et al., 2003).
Germination rate decreased with the increasing of salinity as expected (Table 1). When the two species were compared, the effect of salinity on S. corniculata was higher than that on S. salsa (Table 1). The recovery experiment showed that seeds exposed to different NaCl, NaHCO3 and seawater concentrations for a short period (10 days) could germinate after being transferred to distilled water (Table 1). This is supported by Ungar (1995), who reported that tolerance of seeds to salinity should be interpreted at two levels: (1) the ability to germinate at saline conditions and (2) the ability to germinate at a non-saline condition after exposure to high salinity. In this study the two levels of tolerance of seeds to salinity were observed.
The effects of 3 types of salts on physiological indices: Halophytes were reported to accumulate large amounts of Na+ under salt stress and simultaneously inhibit K+ absorption, which results in the increase of Na+ content and the decrease of K+ content (Shi and Wang, 2005; Yang et al., 2007). Similar phenomenon is observed in this study, results shown in Fig. 1a and c indicated that with increasing stress intensity the Na+ content in seedlings increased, while K+ content and K+/Na+ ratio decreased under NaCl and NaHCO3 stress, but the K+/Na+ ratio were not significantly different among 100, 200 and 300 mM NaCl and NaHCO3 treatments (Fig. 1e). Total accumulation of Na+ and K+ was higher under NaCl stress than that under NaHCO3 stress for both species. This indicated that the negative effects of HCO3- were higher than Cl-. In seawater treatment, the K+ content increased with the increasing of seawater (Fig. 1d), due to the presence of K+ in seawater.
ACKNOWLEDGMENTS
Authors are grateful for support from the Chinese Academy of Sciences (Grant No. kzcx2-yw-223); the National Natural Science Foundation of China ( 30770412 and 40873062); the National Science and Technology Supporting Program of China (Grant No. 2006BAC01A13); the CAS/SAFEA international partnership program for creation research team; the 100 Talents Program of the Chinese Academy of Sciences and the Science and Technology Planning Program of Shandong Province (No. 2008GG20005006, 2008GG3NS07005 and 2005GD42060003).
REFERENCES
- Chen, Z., M. Zhou, I. Newman, N. Mendham, G. Zhang and S. Shabala, 2007. Potassium and sodium relations in salinised barley tissues as a basis of differential salt tolerance. Funct. Plant Biol., 34: 150-162.
Direct Link - Cuin, T.A., A.J. Miller, S.A. Laurie and R.A. Leigh, 2003. Potassium activities in the cell compartments of salt-grown barley leaves. J. Exp. Bot., 383: 657-661.
Direct Link - Garthwaite, A.J., R. von Bothmer and T.D. Colmer, 2005. Salt tolerance in wild Hordeum species is associated with restricted entry of Na+ and Cl-into the shoots. J. Expt. Bot., 56: 2365-2378.
Direct Link - Guan, B., D. Zhou, H. Zhang, Y. Tian, W. Japhet and P. Wang, 2009. Germination responses of Medicago ruthenica seeds to salinity, alkalinity and temperature. J. Arid Environ., 73: 135-138.
CrossRefDirect Link - Keiffer, C.H. and I.A. Ungar, 1997. The effect of extended exposure to hypersaline conditions on the germination of five inland Halophyte species. Am. J. Bot., 84: 104-111.
Direct Link - Khan, M.A. and B. Gul, 1998. High salt tolerance in germinating dimorphic seeds of Arthrocnemum indicum. Int. J. Plant Sci., 159: 826-832.
Direct Link - Khan, M.A., B. Gul and D.J. Weber, 2000. Germination response of Salicornia rubra to temperature and salinity. J. Arid Environ., 45: 207-214.
CrossRef - Khan, M.A. and S. Gulzar, 2003. Germination responses of Sporobolus ioclados: A saline desert grass. J. Arid Environ., 53: 387-394.
CrossRefDirect Link - Khan, M.A. and I.A. Ungar, 1984. Seed polymorphism and germination responses to salinity stress in Atriplex triangularis willd. Bot. Gaz., 145: 487-494.
Direct Link - Mokhberdoran, F., S.M. Nabavi Kalat and R. Sadrabadi Haghighi, 2009. Effect of temperature, iso-osmotic concentrations of NaCl and PEG agents on germination and some seedling growth yield components in rice (Oryza sativa L.). Asian J. Plant Sci., 8: 409-416.
CrossRefDirect Link - Heidari, M., 2009. Variation in seed germination, seedling growth, nucleic acid and biochemical component in canola (Brassica nupus L.) under salinity stress. Asian J. Plant Sci., 8: 557-561.
CrossRefDirect Link - Munns, R., 2005. Genes and salt tolerance: Bringing them together. New Phytol., 167: 645-663.
CrossRefPubMedDirect Link - Munns, R. and R.A. James, 2003. Screening methods for salinity tolerance: A case study with tetraploid wheat. Plant Soil, 253: 201-218.
CrossRef - Noe, G.B. and J.B. Zedler, 2000. Differential effects of four abiotic factors on the germination of salt marsh annuals. Am. J. Bot., 87: 1679-1692.
Direct Link - Ravindran, K.C., K. Venkatesan, V. Balakrishnan, K.P. Chellappan and T. Balasubramanian, 2007. Restoration of saline land by halophytes for Indian soils. Soil Biol. Biochem., 39: 2661-2664.
CrossRefDirect Link - Song, J., H. Fan, Y.Y. Zhao, Y.H. Jia, X.H. Du and B.S.Wang, 2008. Effect of salinity on germination, seedling emergence, seedling growth and ion accumulation of a euhalophyte Suaeda salsa in an intertidal zone and on saline inland. Aquat. Bot., 88: 331-337.
CrossRef - Wang, B.S., U. Luttge and R. Ratajczak, 2004. Specific regulation of SOD isoforms by NaCl and osmotic stress in leavrs of the C3 halophyte Suaeda salsa L. J. Plant Physiol., 161: 285-293.
CrossRef - Wang, Y.C., C.P. Yang, G.F. Liu, G.D. Zhang and Q.Y. Ban, 2007. Microarray and suppression subtractive hybridization analyses of gene expression in Puccinellia tenuiflora after exposure to NaHCO3. Plant Sci., 173: 309-320.
CrossRef - Yang, C.W., J.N. Chong, C.Y. Li, C.M. Kim, D.C. Shi and D. Wang, 2007. Osmotic adjustment and ion balance traits of an alkali resistant halophyte Kochia sieversiana during adaptation to salt and alkali conditions. Plant Soil, 294: 263-276.
CrossRef - Yokoishi, T. and S. Tanimoto, 1994. Seed germination of the halophyte Suaeda japonica under salt stress. J. Plant Res., 107: 385-388.
CrossRef - Zhao, K.F., H. Fan, X.Y. Jiang and S. Zhou, 2002. Critical day-length and photoinductive cycles for the induction of flowering in halophyte Suaeda salsa. Plant Sci., 162: 27-31.
CrossRef - Shi, D. and D. Wang, 2005. Effects of various salt-alkaline mixed stresses on Aneurolepidium chinense (Trin.) Kitag. Plant Soil, 271: 15-26.
CrossRefDirect Link