
Ali Vaziri Gohar
Department of Biochemistry, Afzalipour School of Medicine, Kerman University of Medical Sciences, Kerman, Iran
Abbas Mohammadi
Department of Biochemistry, Afzalipour School of Medicine, Kerman University of Medical Sciences, Kerman, Iran
Fariba Sharififar
Department of Pharmacognosy, School of Pharmacy, Kerman University of Medical Sciences, Kerman, Iran
Asian Journal of Plant Sciences
Year: 2010 | Volume: 9 | Issue: 3 | Page No.: 134-139







ABSTRACT
P53 tumor suppressor protein is a key component of genome guard. The p53 inactivation occurs frequently in many human cancers by mutation or over-expression of its specific inhibitor named MDM2 oncoprotein (Murine Double Minute-2), thus MDM2 inhibitors might offer the preventive and therapeutic strategies. In contrast, ATM (Ataxia Telangiectasia Mutated) tumor suppressor protein activates p53 protein. The MDM2 inhibitors such as Nutlin-3, HLI98 and MDM2 antisense oligonucleotides have been identified, but clinical validation is still pending. The Zataria multiflora Boiss. has antioxidant and antibacterial effects. To determine the correlation between the Zataria multiflora Boiss. and anti-carcinogenesis effects, we investigated the role of the essential oil of this plant on MDM2 and ATM genes expression using semi-quantitative RT-PCR in liver samples of 8 normal male rats. The results clearly showed that ATM and MDM2 gene expression increased and decreased, respectively, although the difference in ATM mRNA between target and control groups did not reach levels of significance. Our data implicated the essential oil of Zataria multiflora Boiss. as a down-regulator of MDM2 gene expression and may offer new tools for therapeutic intervention in the p53 pathway for cancer prevention and treatment.
PDF Abstract XML References Citation
Received: March 02, 2010;
Accepted: April 15, 2010;
Published: June 02, 2010
How to cite this article
Ali Vaziri Gohar, Abbas Mohammadi and Fariba Sharififar, 2010. Role of Zataria multiflora Boiss. Essential oil in Regulation of MDM2 and ATM Genes Expression in Rat. Asian Journal of Plant Sciences, 9: 134-139.
DOI: 10.3923/ajps.2010.134.139
URL: https://scialert.net/abstract/?doi=ajps.2010.134.139
DOI: 10.3923/ajps.2010.134.139
URL: https://scialert.net/abstract/?doi=ajps.2010.134.139
INTRODUCTION
The Zataria multiflora Boiss. with the local name of Avishan-e-Shirazi in Iran is a plant belonging to the Laminaceae family that grows only in Iran, Pakistan and Afghanistan (Ali et al., 2000). It has been used for several traditional purposes as a Xavor ingredient and natural preservative in a wide variety of food. Also, it has been used for medicinal purposes including pre-mature labor pains, rupture, antiseptic, anaesthetic, antispasmodic and antinociceptive activity on Salmonella typhimurium and Staphylococcus aureus in brain heart infusion broth (Hossinzadeh et al., 2000; Ebrahimzadeh et al., 2003; Fazeli et al., 2007; Misaghi and Basti, 2007; Sharififar et al., 2007).
Essential oils are naturally occurring terpenic mixtures isolated from various parts of plants (Daferera et al., 2000). The twenty-five compounds were identified in the essential oil of this plant. The main constituents are phenolic compounds such as thymol and carvacrol that consist of 37.7 and 33.6%, respectively. The other compounds are p-cymene, c-terpinene, b-caryophyllene, b-caryophyllene and monoterpenes (Shafiee and Javidnia, 1997; Ebrahimzadeh et al., 2003; Sharififar et al., 2007). Thymol and arvacrol have antioxidant properties and are able to inhibit linoleic oxidation that were detected by using two separate methods including inhibition of free radical 2,2-diphenyl-1-picrylhydrazyl (DPPH) and ammonium thiocyanate systems (Sharififar et al., 2007).
TP53 tumor suppressor gene encodes a protein as a guard of genome that has a key role in the control of cell cycle, cell differentiation and apoptosis (Ng et al., 1999; Mohammadi et al., 2008). Upon DNA damage, a complex network of responses is initiated. The p53 protein with its leading role stabilized through post-translational modifications including phosphorylation and acetylation in the nucleus to initiate the transcription of downstream genes that are responsible for cell cycle arrest in G1 and G2 checkpoints or apoptosis (Ashcroft and Vousden, 1999; Liu et al., 2004).
P53 protein activity is either disabled or attenuated by TP53 gene mutation, loss of heterozygosity (LOH) of 17p13 band and higher activity of the MDM2 (Murine Double Minute 2) oncoprotein as a best-characterized negative regulator of the p53 protein (Toledo and Wahl, 2007).
MDM2 proto-oncogene was discovered as the second gene in the amplified region of the double minute chromosome in the spontaneously transformed 3T3-DM mouse cell line (Cahilly-Snyder et al., 1987; Fakharzadeh et al., 1991). This proto-oncogene leads to a prominent oncogene through gene amplification. The MDM2 oncogene encodes a 90 KDa nuclear phosphoprotein act as the E3 ubiquitin ligase with three conserved domains including N-terminal domain (BoxI Binding Domain as the most conserved domain), Zinc finger domain and C-terminal RING domain (Liu et al., 2004; Toledo and Wahl, 2007). This protein binds with its N-terminal domain to only three amino acids of N-terminal domain (transactivation domain) of p53 protein. This event leads to two different pathways including rapid ubiquitin-dependent proteasomal degradation of p53 protein and inhibition of the interaction with its transcriptional co-activators such as p300 and CBP (CREB-Binding Protein). Therefore, destabilization of MDM2 oncoprotein is essential for proper p53 function (Maki et al., 1996; Kubbutat et al., 1997).
Levels and stability of MDM2 are also modulated by ubiquitylation (Honda and Yasuda, 2000). MDM2 also interacts with an ubiquitin specific protease USP7, which can reverse MDM2-ubiquilyation and prevent it from being degradation. Also, it is interesting to note that USP7 protects p53 protein from degradation (Meulmeester et al., 2005; Sheng et al., 2006).
In contrast to MDM2 oncoprotein, the ATM tumor suppressor protein is positive regulator of p53 protein. The product of ATM gene is a serine/threonine-specific protein kinase with EC No. 2.7.11.1 belonging to the PI3/PI4-kinase family that recruited and activated by DNA double-strand breaks (DSBs). ATM phosphorylates and activates several key proteins including several checkpoint proteins p53, BRCA1, CHK2, RAD17 and RAD9 leading to cell cycle arrest, DNA repair or finally apoptosis (Banin et al., 1998; Goodarzi and Lees-Miller, 2004).
The p53 activation through ATM results from three different pathways. First, ATM phosphorylates the ser15 of p53 protein leads to reduce interaction between p53 and MDM2. Second, ATM directly phosphorylates Ser395 on MDM2, which impairs nuclear export and degradation of p53 finally ATM mediates indirect phosphorylation of MDM2 on Tyr394 via the c-Abl kinase. Follow Ser15 phosphorylation, p53 is phosphorylated on Thr18, which might affect the MDM2-p53 interaction. The ATR (ATM-related kinase) maintains phosphorylation of Ser15 at later stages during the DNA damage response.
The goal of this work is to identify the Zataria multiflora Boiss. properties on gene expression of two key genes related to cell cycle control.
MATERIALS AND METHODS
Plant material:
The tops at the full flowering stage (June and July, 2009) were collected from plants growing wild in the Firoozabad, Fars, Iran. The taxonomic identification of plant materials was confirmed by a senior plant taxonomist. Voucher specimens were deposited in the Herbarium of Kerman Faculty of Pharmacy, Kerman, Iran.
Isolation of the essential oil:
The air-dried and ground herbal parts of the plant collected were submitted for 4 h to water-distillation using a British-type Clevenger apparatus (yield 2.8% v/w). The obtained essential oil was dried over anhydrous sodium sulphate then stored at +4°C until tested and analyzed (Sharififar et al., 2007).
Animal groups:
Eight male Sprague-Dawley rats (6 weeks old) were obtained from animal room of Afzalipour School of Medicine, Kerman University of Medical Sciences, Kerman, Iran. The rats in two groups (control and essential oil+) were randomly assigned to semisynthetic diets based on torula yeast, corn starch. The rats were maintained under standard conditions at 22°C, with 12:12-h dark/light cycles in store room since two weeks before treatment. They consumed the diets ad libitum and had free access to water.
Tests preparation:
Dimethyl sulphoxide (400 μL/kg/day) and essential oil of Zataria multiflora Boiss. (50 μL/kg/day dissolved in 400 μL/kg/day Dimethyl sulphoxide) were injected intraperitoneally (IP) for 7 days at light cycle for control and essential oil rat groups, respectively. This concentration of essential oil is the highest dose that is not lethal in rat. After one week rats were sacrificed by decapitation. Then, liver samples were rapidly removed and individual sections was placed under liquid nitrogen then stored at -70°C for subsequent analysis. Our animal protocol was approved by Vice Chancellor for Research, Kerman University of Medical Sciences, Kerman, Iran.
RNA extraction:
All pipette tips and tubes were treated with 0.1% diethyl pyrocarbonate (DEPC) solution at 37°C for overnight, then heated at 100°C for 30 min and finally autoclaved. Between 25-50 mg liver tissue was lysised by sonicator on ice. Total RNA was prepared using RNA tissue extraction kit (Roche Applied Sciences, Germany) based on manual protocol and extracted RNA solved in DEPC-treated water for further processing. In this procedure, Dnase I enzyme (Roche Applied Sciences, Germany) was used to eliminate the DNA source of samples. The concentration of total RNA was measured by spectrophotometer NanoDrop ND-1000 and finally, the tubes stored at -80°C.
RT-PCR reaction:
cDNA was synthesized using First Strand cDNA synthesis (Fermentas, litany) in 25 μL volume, was utilized 1 μg total RNA, 1 μL oligo-dT(18), 4 μL buffer, 1 μL RNase inhibitor enzyme, 2 μL dNTPs mix (10 mM) and DEPC-treated water and finally 5 U of M-MuLV Reverse Transcriptase (Fermentas, Litany). Then second strand was made by Rnase H (Fermentas, Litany) and DNAPI (Fermentas, Litany). The sequence of primers and NCBI Reference numbers for cDNA fragments amplification were: ATM, 5-AAG-CGG-GTG-TGT-GAA-ATA-GG-3 (forward) and 5-AGT-GGA-TTC-CCT-TTG-TGT-CG-3 (reverse) with NM_001106821.1; MDM2: 5-CTT-CGT-GAG-AAC-TGG-CTT-CC-3 (forward) and 5-AAT-CAT-TTG-GAT-CGG-CTG-TC-3 (reverse) with NM_001108099.1; Beta-actin as the control gene, 5-TCG-TGG-GCC-GCC-CTA-GGC-AC-3 (forward) and 5-GGC-CTT-AGG-GTT-CAG-AGG-GGC-3 (reverse) with NM_031144.2. The Primers were synthesized by Eurofins MWG Operon, Germany. The semi-quantitative RT-PCR was performed in triplicate using Termal Cycler (Corbett, Australia) in final volume of 25 μL reaction mixture containing: 1x PCR Buffer (Fermentas, Litany), 0.2 mM dNTP mix (Fermentas, Litany), 2 mM MgCl2 (Fermentas, Litany), o.5 μM each forward and reverse primers, 3 U of Taq DNA polymerase (Fermentas, Litany), cDNA and RNase-free water (Fermentas, Litany). The mixture was heated at 95°C for 3 min and then subjected to 40 cycles of PCR as follows: 94°C for 40 sec, 60°C for 40 sec, 72°C for 40 sec and final extension at 72°C for 5 min. The PCR product lengths of ATM, MDM2 and beta-actin genes were 387, 480 and 256 bp. Then, PCR products were loaded on a 2% gel agarose electrophoresis. All tests were performed in triplicate and then, the intensity (OD) of the bands was measured by image-i software (version 3.0) in OD target gene/OD control gene.
Statistical analysis:
Data are presented as mean±standard error of the mean (S.E.M). Statistical analysis was carried out by independent-sample t-test (SPSS for Windows version 11.5). Differences were considered significant if p<0.05.
RESULTS AND DISCUSSION
Expression of ATM tumor suppressor gene:
To determine the effects of essential oil of Zataria multiflora Boiss. on gene expression, we measured mRNA expression by semi-quantitative RT-PCR in eight male rats. As shown in Fig. 1a-c, treatment of essential oil of Zataria multiflora Boiss. caused a 1.6-fold increase in ATM mRNA level compared to control group, although this difference did not reach levels of significance (p = 0.24).
Expression of MDM2 oncogene:
To determine the effects of essential oil of Zataria multiflora Boiss. on gene expression, we measured mRNA expression by semi-quantitative RT-PCR in eight male rats. As shown in Fig. 2a-c, treatment of essential oil of Zataria multiflora Boiss. caused approximately 2-fold significantly decrease in MDM2 mRNA level compared to control group (p = 0.001).
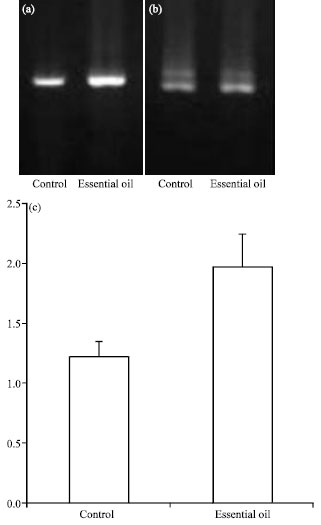 | |
| Fig. 1: | The effect of Zaratia multiflora Boiss. on ATM tumor suppressor gene expression. (a) Two percent agarose gel electrophoresis of RT-PCR products of ATM gene from control and essential oil-treated groups. (b) Two percent agarose gel electrophoresis of RT-PCR products of Beta-actin as control gene from control and essential oil-treated groups. (c) Treatment of Zaratia multiflora Boiss. caused a 1.6-fold increase in ATM mRNA level compared to control group, although this difference did not reach levels of significance. The arrow indicates SEM |
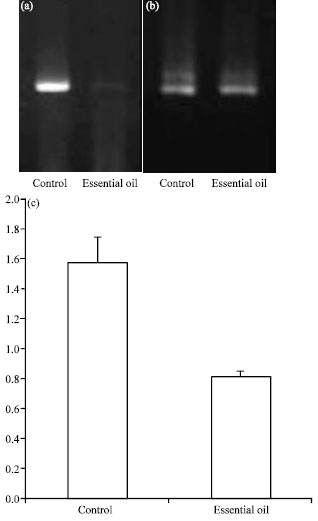 | |
| Fig. 2: | The effects of Zaratia multiflora Boiss. on MDM2 oncogene expression. (a) Two percent agarose gel electrophoresis of RT-PCR products of MDM2 gene from control and essential oil-treated groups. (b) Two percent agarose gel electrophoresis of RT-PCR products of Beta-actin as control gene from control and essential oil-treated groups. (c) Treatment of Zaratia multiflora Boiss. caused about 2-fold significant increase in MDM2 mRNA level compared to control group. The arrow indicates SEM |
As p53 protein has a key role in the cellular defence mechanisms against neoplastic transformation, thus p53 regulators such as ATM and MDM2 proteins have so important role in cell cycle regulation. In normal cells, p53 and MDM2 participate in an autoregulatory loop in which p53 up-regulates MDM2 expression MDM2 induce p53 degradation by the proteasome (Chen et al., 1998; Bauer et al., 2010).
The TP53 gene inactivation in some human cancers including soft tissue sarcomas, osteosarcomas, breast tumors, retinoblastoma and neuroblastoma cell lines is not related to gene mutation or deletion, but is results from MDM2 amplification (Momand et al., 1998; Lowe and Sherr, 2003; Michael and Oren, 2003; Sherr, 2006). Also, ATM gene inactivation induces 100-fold increased of cancer types (Kurz and Lees-Miller, 2004; Halaby et al., 2008). Therefore, identification of suitable MDM2 inhibitors and ATM inducers has been proposed as a therapeutic strategy for a large population of cancer patients.
Since the essential oils of medicinal plants are eco-friendly and harmless to humans, there is an increasing attention for usage of these compounds in industry, academic research and medical (Paranagama et al., 2003; Velluti et al., 2003; Singh et al., 2006).
The present study displayed that the essential oil of Zataria multiflora Boiss. has both suppressing MDM2 and inducing ATM gene expression effects, although the later was not significance. However, the fraction of essential oil that has these potential effects remains an important issue to resolve. Also, the results indicated that the basal expression of MDM2 gene was higher than the basal expression of ATM gene in rat liver, although this difference did not reach levels of significance (p = 0.06).
There are some natural MDM2 inhibitors including p14ARF, PI3KKs, ATR and Akt/PKB in vivo. The p14ARF kinase is an inhibitor of MDM2 ubiquitin ligase activity. DNA damage induces PI3K kinase directly phosphorylates MDM2 protein on Ser395 and destabilizes MDM2 protein temporarily. The ATR kinase directly phosphorylates MDM2 protein on Ser407 in response to replication block. The Akt/PKB regulates the intracellular localization of MDM2 by phosphorylation on serines 166 and 186. These pathways can reduce the ability of MDM2 to degrade p53 after DNA damage (Banin et al., 1998; Honda and Yasuda, 1999; Abraham, 2001; Zhou et al., 2001; Yang et al., 2003; Koundrioukoff et al., 2004; Vaziri Gohar et al., 2007).
Also, several MDM2 antagonists have been synthesized such as Nutlin-3, HLI98 and MDM2 antisense oligonucleotides. Nutlin-3 is a cis-imidazole analog that mimic the peptides of BoxI binding domain of MDM2 protein leads to disrupt p53-MDM2 interactions, in vitro. Oral administration of nutlin-3 reduced tumor growth and metastasis in the presence of wild-type p53. HLI98 specifically targets the E3 ligase activity, thus inhibits MDM2-dependent p53 degradation, but was not tested in vivo. MDM2 antisense oligonucleotide as a sequence-specific post translational gene silencing mechanism inhibits MDM2 expression in cells and mouse models of human cancer resulted in p53 stabilization not only wild-type p53 cells, but also in mutant-type p53 cells (Tortora et al., 2000; Zhang et al., 2004, 2005; Yang et al., 2005; Vassilev, 2007; Toledo and Wahl, 2007).
Also, treatment of SW480 cells with Ras inhibitor FTS (farnesylthiosalicylic acid) caused a distinct decrease in MDM2 oncogene and caused a 1.6-fold increase in p53 mRNA (Halaschek-Wiener et al., 2004).
The present study showed the essential oil of Zataria multiflora Boiss. has anti-carcinogenesis effects by MDM2 oncogene attenuation and ATM tumor suppressor gene activation in rat liver and therefore provides more evidences for its preventive and therapeutic application on cancer therapy.
ACKNOWLEDGMENTS
This project is supported by grant No. 87/10/60/1301 of Vice Chancellor for Research, Kerman University of Medical Sciences, Kerman, Iran. Also, authors are grateful to Dr. Gholamhoseinian Najar for his kindness in providing us with some devices and materials.
REFERENCES
- Abraham, R.T., 2001. Cell cycle checkpoint signaling through the ATM and ATR kinases. Genes Dev., 15: 2177-2196.
Direct Link - Ali, M.S., M. Saleem, Z. Ali and V.U. Ahmad, 2000. Chemistry of Zataria Multiflora (Lamiaceae). Phytochemistry, 55: 933-936.
CrossRef - Ashcroft, M. and K.H. Vousden, 1999. Regulation of p53 stability. Oncogene, 18: 7637-7643.
Direct Link - Banin, S., L. Moyal, S. Shieh, Y. Taya and C.W. Anderson et al., 1998. Enhanced phosphorylation of p53 by ATM in response to DNA damage. Science, 281: 1674-1677.
Direct Link - Bauer, S., T. Muhlenberg, M. Leahy, M. Hoiczyk, T. Gauler, M. Schuler and L. Looijenga, 2010. Therapeutic potential of Mdm2 inhibition in malignant germ cell tumours. Eur. Urol., 57: 679-687.
CrossRef - Cahilly-Snyder, L., T. Yang-Feng, U. Francke and D.L. George, 1987. Molecular analysis and chromosomal mapping of amplified genes isolated from a transformed mouse 3T3 cell line. Somat. Cell Mol. Genet., 13: 235-244.
CrossRef - Chen, L., S. Agrawal, W. Zhou, R. Zhang and J. Chen, 1998. Synergistic activation of p53 by inhibition of MDM2 expression and DNA damage. Proc. Natl. Acad. Sci., 95: 195-200.
Direct Link - Daferera, D.J., B.N. Ziogas and M.G. Polissiou, 2000. GC-MS analysis of essential oils from some Greek aromatic plants and their fungitoxicity on Penicillium digitatum. J. Agric. Food Chem., 48: 2576-2581.
CrossRefDirect Link - Ebrahimzadeh, H., Y. Yamini, F. Sefidkon, M. Chaloosi and S.M. Pourmortazavi, 2003. Chemical composition of the essential oil and supercritical CO2 extracts of Zataria multiflora Boiss. Food Chem., 83: 357-361.
Direct Link - Fakharzadeh, S.S., S.P. Trusko and D.L. George, 1991. Tumorigenic potential associated with enhanced expression of a gene that is amplified in a mouse tumor cell line. EMBO J., 10: 1565-1569.
Direct Link - Fazeli, M.R., G. Amin, M.M.A. Attari, H. Ashtiani, H. Jamalifar and N. Samadi, 2007. Antimicrobial activities of Iranian sumac and avishan-e shirazi (Zataria multiflora) against some food-borne bacteria. Food Control, 18: 646-649.
CrossRefDirect Link - Goodarzi, A.A. and S.P. Lees-Miller, 2004. Biochemical characterization of the ataxia-telangiectasia mutated (ATM) protein from human cells. DNA Repair, 3: 753-767.
Direct Link - Halaby, M.J., J.C. Hibma, J. He and D.Q. Yang, 2008. ATM protein kinase mediates full activation of Akt and regulates glucose transporter 4 translocation by insulin in muscle cells. Cell Signal., 20: 1555-1563.
Direct Link - Halaschek-Wiener, J., V. Wacheck, Y. Kloog and B. Jansen, 2004. Ras inhibition leads to transcriptional activation of p53 and down-regulation of MDM2: two mechanisms that cooperatively increase p53 function in colon cancer cells. Cell Signal., 16: 1319-1327.
Direct Link - Honda, R. and H. Yasuda, 1999. Association of p19(ARF) with Mdm2 inhibits ubiquitin ligase activity of Mdm2 for tumor suppressor p53. EMBO J., 18: 22-27.
Direct Link - Honda, R. and H. Yasuda, 2000. Activity of MDM2, a ubiquitin ligase, toward p53 or itself is dependent on the RING finger domain of the ligase. Oncogene, 19: 1473-1476.
Direct Link - Hosseinzadeh, H., M. Ramezani and G.A. Salmani, 2000. Antinociceptive, anti-inflammatory and acute toxicity effects of Zataria multiflora boiss extracts in mice and rats. J. Ethnopharmacol., 73: 379-385.
CrossRefPubMedDirect Link - Koundrioukoff, S., S. Polo and G. Almouzni, 2004. Interplay between chromatin and cell cycle checkpoints in the context of ATR/ATM-dependent checkpoints. DNA Repair, 3: 969-978.
Direct Link - Kubbutat, M.H.G., S.N. Jones and K.H. Vousden, 1997. Regulation of p53 stability by Mdm2. Nature, 387: 299-303.
Direct Link - Kurz, E.U. and S.P. Lees-Miller, 2004. DNA damage-induced activation of ATM and ATM-dependent signaling pathways. DNA Repair, 3: 889-900.
CrossRef - Liu, Z., E.T. Olejniczak and S.W. Fesik, 2004. Over-expression of the human MDM2 p53 binding domain by fusion to a p53 transactivation peptide. Protein Exp. Purif., 37: 493-498.
Direct Link - Lowe, S.W. and C.J. Sherr, 2003. Tumor suppression by Ink4a-Arf: Progress and puzzles. Curr. Opin. Genet. Dev., 13: 77-83.
Direct Link - Maki, C.G., J.M. Huibregtse and P.M. Howley, 1996. In vivo ubiquitination and proteasome-mediated degradation of p53(1). Cancer Res., 56: 2649-2654.
Direct Link - Meulmeester, E., Y. Pereg, Y. Shiloh and A.G. Jochemsen, 2005. ATM-mediated phosphorylations inhibit mdmx/MDM2 stabilization by HAUSP in favor of p53 activation. Cell Cycle, 4: 1166-1170.
Direct Link - Michael, D. and M. Oren, 2003. The p53-Mdm2 module and the ubiquitin system. Semin Cancer Biol., 13: 49-58.
CrossRefPubMedDirect Link - Misaghi, A. and A.A. Basti, 2007. Effect of Zataria Multiflora Boiss. essential oil and nisin on Bacillus cereus ATCC 11778. Food Control, 18: 1043-1049.
CrossRef - Mohammadi, A., A. Vaziri Gohar and M.R. Shakibaie, 2008. Mutations in tumor suppressor TP53 gene in formalin- fixed, paraffin embedded tissues of Squamous Cell Carcinoma (SCC) of lung cancer. Am. J. Biochem. Biotehcnol., 4: 1-6.
CrossRefDirect Link - Momand, J., D. Jung, S. Wilczynski and J. Niland, 1998. The MDM2 gene amplification database. Nucl. Acids Res., 26: 3453-3459.
Direct Link - Ng, I.O.L., K.Y. Lam, M. Ng and J.A. Regezi, 1999. Expression of p21/waf1 in oral squamous cell carcinomas-correlation with p53 and MDM2 and cellular proliferation index. Oral Oncol., 35: 63-69.
Direct Link - Paranagama, P.A., K.H.T. Abeysekera, K. Abeywickrama and L. Nugaliyadde, 2003. Fungicidal and anti-aflatoxigenic effects of the essential oil of Cymbopogon citratus (DC.) Stapf. (lemongrass) against Aspergillus flavus Link. isolated from stored rice. Lett. Appl. Microbiol., 37: 86-90.
CrossRefPubMedDirect Link - Shafiee, A. and K. Javidnia, 1997. Composition of essential oil of Zataria multiflora. Plant. Media., 63: 371-372.
Direct Link - Sharififar, F., M.H. Moshafi, S.H. Mansouri, M. Khodashenas and M. Khoshnoodi, 2007. In vitro evaluation of antibacterial and antioxidant activities of the essential oil and methanol extract of endemic Zataria multiflora Boiss. Food Control, 18: 800-805.
CrossRefDirect Link - Sheng, Y., V. Saridakis, F. Sarkari, S. Duan and T. Wu et al., 2006. Molecular recognition of p53 and MDM2 by USP7/HAUSP. Nat. Struct. Mol. Biol., 13: 285-291.
CrossRefPubMedDirect Link - Sherr, C.J., 2006. Divorcing ARF and p53: An unsettled case. Nat. Rev. Cancer, 6: 663-673.
Direct Link - Singh, G., S. Maurya, M.P. Lampasona and C. Catalan, 2006. Chemical constituent, antifungal and antioxidative potential of Foeniculum vulgare volatile and its acetone extract. Food Control, 17: 745-752.
CrossRef - Toledo, F. and G.M. Wahl, 2007. MDM2 and MDM4: p53 regulators as targets in anticancer therapy. J. Biochem. Cell Biol., 39: 1476-1482.
Direct Link - Velluti, A., V. Sanchis, A.J. Ramos, J. Egido and S. Marin, 2003. Inhibitory effect of cinnamon, clove, lemongrass, oregano and palmarose essential oils on growth and fumonisin B1 production by Fusarium proliferatum in maize grain. Int. J. Food Microbiol., 89: 145-154.
CrossRefDirect Link - Yang, J., Y. Yu, H.E. Hamrick and P.J. Duerksen-Hughes, 2003. ATM, ATR and DNA-PK: Initiators of the cellular genotoxic stress responses. Carcinogenesis, 24: 1571-1580.
Direct Link - Yang, Y., R. Ludwig, J. Jensen, S. Pierre and M. Medaglia et al., 2005. Small molecule inhibitors of HDM2 ubiquitin ligase activity stabilize and activate p53 in cells. Cancer Cell, 7: 547-559.
Direct Link - Zhang, R., H. Wang and S. Agrawal, 2005. Novel antisense anti-MDM2 mixed-backbone oligonucleotides: proof of principle, in vitro and in vivo activities and mechanisms. Curr. Cancer Drug Targets, 5: 43-49.
Direct Link - Zhang, Z., H. Wang, M. Li, S. Agrawal and X. Chen et al., 2004. MDM2 is a negative regulator of p21WAF1/CIP1, independent of p53. J. Biol. Chem., 279: 16000-16006.
Direct Link - Zhou, B.P., Y. Liao, W. Xia, Y. Zou and B. Spohn et al., 2001. HER-2/neu induces p53 ubiquitination via Akt-mediated MDM2 phosphorylation. Nat. Cell Biol., 3: 973-982.
Direct Link