
H. Tanaka
Frontier Science Research Center, University of Miyazaki, 1-1 Gakuen Kibanadai Nishi, Miyazaki 889-2192, Japan
J. Toyama
Frontier Science Research Center, University of Miyazaki, 1-1 Gakuen Kibanadai Nishi, Miyazaki 889-2192, Japan
R. Akashi
Frontier Science Research Center, University of Miyazaki, 1-1 Gakuen Kibanadai Nishi, Miyazaki 889-2192, Japan
Asian Journal of Plant Sciences
Year: 2009 | Volume: 8 | Issue: 8 | Page No.: 544-550







ABSTRACT
This study examines the function and genetic structure of Momordica charantia lectin. A galactose-binding lectin (MCL1) was purified from M. charantia seeds. The MCL1 showed highest hemagglutinating activity toward human type-O(H) erythrocytes followed by A, B and Omh (para-Bombay phenotype, also known as H-deficient secretor) erythrocytes. Moreover, we observed that MCL1 inhibited the cell-free synthesis of luciferase in a rabbit reticulocyte lysate system. The N-terminal amino acid sequence of purified MCL1 was identified and used to design degenerate oligonucleotide primers. The 3' and 5' ends of the gene coding for this protein were amplified by rapid amplification of cDNA ends, cloned and sequenced. The coding region (1641 bp, 547 amino acid residues) consisted of a 23 amino acid N-terminal signal sequence preceding an A-chain of 263 amino acid residues encoding a ribosome-inactivating protein that was joined to the B-chain of 261 amino acid residues encoding a lectin. The transcript was detected only in embryos, but hemagglutinating activity was detected both in embryos and cotyledons. These results suggest that gene expression occurred only during embryogenesis and the product accumulated in embryos and cotyledons. The MCL1 was expressed in tobacco BY-2 cells and the supernatant fluid of disrupted cells showed higher hemagglutinating activity toward human type-O(H) erythrocyte than the other tested erythrocytes. Thus, transgenic tobacco suspension culture cells harboring the cloned cDNA encoding the lectin purified from M. charantia are expected to be useful for the production of MCL1.
PDF Abstract XML References Citation
How to cite this article
H. Tanaka, J. Toyama and R. Akashi, 2009. Molecular Characterization of a Galactose-Binding Lectin from Momordica
charantia Seeds and Its Expression in Tobacco Cells. Asian Journal of Plant Sciences, 8: 544-550.
DOI: 10.3923/ajps.2009.544.550
URL: https://scialert.net/abstract/?doi=ajps.2009.544.550
DOI: 10.3923/ajps.2009.544.550
URL: https://scialert.net/abstract/?doi=ajps.2009.544.550
INTRODUCTION
Lectins are non-catalytic proteins or glycoproteins that are capable of recognizing and binding reversibly to specific saccharide moieties of glycoconjugates (Goldstein et al., 1980). Lectins have a variety of biological properties including antifungal (Melo et al., 2005), antiproliferative (Kaur et al., 2005) mitogenic (Wong et al., 2006) and hemagglutinating activities. Some lectins possess a domain with a distinct biological activity. Lectins belonging to the group of type-2 Ribosome-Inactivating Proteins (RIP) consist of a carbohydrate binding B-chain linked to an A-chain that has a highly specific rRNA-N-glycosidase activity and are capable of catalytically inactivating ribosomes. Type-2 RIPs have been grossly divided into two groups: toxic and non-toxic, based on the considerable differences of their cytotoxicity and consequences of their toxicity to animals (Stirpe and Battelli, 2006). On the other hand, type-1 RIP consists of a single, catalytically active subunit and has comparatively weaker cytotoxicity than type-2, because type-1 cannot penetrate through cell membranes. All type-1 RIPs tested so far have antiviral activity against plant, fungal and animal viruses, whereas only a few type-2 RIPs have been reported to be active (Chen et al., 2002; Stirpe and Battelli, 2006; Xu et al., 2007).
Momordica charantia (bitter gourd), a non-leguminous climber belonging to the Cucurbitaceae, has been widely used in the Orient as a foodstuff, although the seeds are not consumed. Some lectins from M. charantia have been isolated and characterized (Barbieri et al., 1980; Wang and Ng, 1998; Toyama et al., 2008) and some researchers have suggested that the lectin has RIP activity. Amino acid compositions and/or partial sequences of these lectins have been revealed in previous reports; however, the genes for these lectins have not been cloned and sequenced yet. In the present study, we describe the cloning and sequencing of cDNAs encoding a galactose-binding lectin from the seeds of M. charantia and monitored gene expression in different tissues using reverse transcription PCR (RT-PCR).
MATERIALS AND METHODS
The studies were conducted at University of Miyazaki during the period of 2004-2008.
Sample materials: Momordica charantia [L.] seeds were collected from industrial waste of a food-processing company (Ishihara Foods, Miyazaki, Japan). Fresh embryo (collected from immature fruits) and other tissues were harvested and used immediately or stored at -80°C in RNAlater (Applied Biosystems, Foster City, CA, USA).
Hemagglutination and inhibition of protein synthesis: The hemagglutinating activity of lectin extracts were assayed by the twofold serial-dilution method against 3% human erythrocyte suspensions (Ortho-Clinical Diagnostics, Tokyo, Japan). A nonradioactive assay for inhibition of protein synthesis was performed as described by Langer et al. (1996). Inhibition of protein synthesis was determined quantitatively by measuring luciferase activity in a cell-free rabbit reticulocyte lysate system (Promega, Madison, WI, USA). The luciferase activities were measured using the Luciferase Assay System (Promega) and a microplate reader (GENios; Tecan, Research Triangle Park, NC, USA).
Total RNA isolation and cDNA cloning: Total RNA was isolated and purified from 70 mg of M. charantia embryos using an RNAqueous column with the Plant RNA Isolation Aid (Applied Biosystems) according to the manufacturer's instructions. The 3' and 5' ends of the transcripts were amplified by Rapid Amplification of cDNA Ends (RACE) method using a GeneRacer Kit (Invitrogen, Carlsbad, CA, USA). For 3' RACE, the forward primer P1 (5'-AAYGARCARTGYTCWCC-3') corresponding to the N-terminal amino acid sequence NEQCSP and the GeneRacer 3' primer provided in the kit were used. For 5' RACE, the GeneRacer 5' primer provided in the kit and a reverse primer P2 (5'-CATGCATTGAGTTCATGTGTGGATAAGC-3') were used. The 5' and 3' RACE products were cloned into pCR4-TOPO with a TOPO TA-Cloning kit (Invitrogen) and then sequenced. Sequence data were analyzed using GENETYX-MAC software package (Software Development, Tokyo, Japan). Phylogenetic trees were constructed from the aligned amino acid sequences using the MEGA software package (Kumar et al., 2004).
Expression of M. charantia MCL1 in tobacco BY-2 cells: MCL1 cDNA was expressed in tobacco BY-2 suspension cells (Nicotiana tabacum L. cv. Bright Yellow-2) (Kato et al., 1991) using the binary vector pH35CG (Inplanta Innovations, Yokohama, Japan) introduced by the particle bombardment method (Finer et al., 1992; Takeuchi et al., 1992; Akashi et al., 2002). The vector contains MCL1 cDNA linked to the cauliflower mosaic virus 35S promoter and the nopaline synthase (NOS) terminator. A hygromycin phosphotransferase gene, conferring hygromycine tolerance, is linked to the NOS promoter. The vector was coated onto gold particles (1.5-3 μm diameter; Aldrich Chemical, Milwaukee, WI, USA) following the protocol described previously (Akashi et al., 2002; Gondo et al., 2005). Bombardment was carried out at a reduced air pressure of -0.1 MPa, target distance of 9.6 cm, helium pressures of 5 kg cm-2 and single shot per plate-mode. Bombarded cells were placed on solidified LS medium (Linsmaier and Skoog, 1965) containing 50 mg mL-1 hygromycin and subsequently subcultured several times for 14 day periods on the same medium in the dark. After 50 day of subculture under selective conditions, hygromycin-resistant cells were grown on a rotary shaker (110 rpm) in 500 mL Erlenmeyer flasks containing 100 mL of LS medium at 27°C in the dark for 10 day. The suspended cells were collected and disrupted by using a Tissue Ruptor (Qiagen, Valencia, CA, USA) on ice. The disrupted cells were centrifuged at 20,000x g for 10 min. The supernatant fluids were assayed for hemagglutinating activities as described earlier.
Expression analysis: The expression of MCL1 mRNA in different tissues was measured by RT-PCR. Tissues, roots, shoots, leaves, cotyledons and embryos of M. charantia were collected and total RNA was extracted using TRIzol Reagent (Invitrogen). The first-strand cDNA was synthesized using a QuantiTect Reverse Transcription kit (Qiagen) according to the manufacturer's instructions. The analysis was performed based on a 99 bp amplicon generated using gene-specific primers P3 (5'-TTTGCGAACGCTTCCTTCTC-3') and P4 (5'-CACGATATGCTGCGAATCCA-3'). The expression of a housekeeping 18S rRNA of M. charantia (GenBank accession number AY900000) as an endogenous reference was carried out using a set of 18S rRNA primers (5'-CTCCGGCGCTGTTACTTTGA-3' and 5'-TCCCGAAGGCCAACAGAATA-3'). The thermal cycling program consisted of an initial step at 94°C for 60 sec, followed by 50 cycles of 94°C for 30 sec and 60°C for 30 sec. The PCR products were separated in 2% agarose gel and visualized by ENVISION DNA dye as a loading buffer (Amresco, Solon, OH, USA).
Nucleotide sequence accession number: The nucleotide sequence of the 1,698-bp region containing MCL1 gene will appear in the DDBJ/EMBL/GenBank nucleotide sequence databases with the accession number AB373132.
RESULTS AND DISCUSSION
Purification and N-terminal amino acid sequences of MCL1 subunits: The anti-H lectin was purified from M. charantia seeds as described by Toyama et al. (2008). The protein showed higher hemagglutinating activity toward human type-O erythrocyte followed by -A2, -A1, -B and -Omh (para-Bombay phenotype, also known as H-deficient secretor) erythrocytes. These observations indicated that the purified protein has anti-H activity. In a previous study, anti-H lectin was dissociated into four subunits, designated MCL1-a, -b, -c and -d, under reducing conditions (Toyama et al., 2008). N-terminal amino acid sequencing identified the first 8 to 20 amino acid residues of each subunit: MCL1-a, NEQCSPQQRTT; MCL1-b, NEQCSPQQ; MCL1-c, NLSLSQSXFSADTYKSFIKN; and MCL1-d, NLSLSQS.
Isolation and characterization of the gene encoding MCL1: We have isolated cDNA clones encoding a precursor protein (MCL1) of 547 amino acid residues with a calculated molecular weight of 60,993 Da (Fig. 1). The N-terminal amino acid sequences of the mature protein purified from seeds of M. charantia matched with the deduced amino acid sequences of Asn-24 to Asn-30 of MCL1-c and/or -d and Asn-287 to Thr-297 of MCL1-a and/or -b. Wang and Ng (1998) purified a lectin from M. charantia and identified the sequence of the first 50 N-terminal amino acids.
 | |
| Fig. 1: | Nucleotide sequence of the MCL1 gene and deduced amino acid sequence. The non-coding sequences are shown as lower-case letters. Amino acid sequences underlined by a thick line are identical to those found in the purified protein. Potential sites for N-linked glycosylation are underlined. Five QxW repeats are double underlined. The positions of gene-specific primers, P1 to P4, are indicated by horizontal arrows above the sequences |
The first 50 residues share 76% sequence similarity with MCL1, suggesting that the two proteins are different. According to the described method of von Heijne (1986) for predicting signal sequence cleavage sites, the signal peptide of MCL1 was most probably cleaved between Cys-23 and Asn-24, a finding in agreement with the N-terminal amino acid sequences of MCL1-c and -d of this study as described above. Proteolytic processing of this polypeptide showed the A- and B-chains of mature MCL1 and implicated more proteolytic cleavages. Since MCL1-a and -b started with the sequence NEQCSPQQ as described above, cleavage must have taken place between Asn-286 and Asn-287 of the precursor after cleavage of the signal peptide to yield an A-chain of 263 amino acid residues encoding a RIP domain with a calculated molecular weight of 29,473 and a B-chain of 261 amino acid residues encoding a lectin domain with a calculated molecular weight of 28,991. To determine if protein synthesis could be inhibited by the purified lectin, luciferase activity was measured in the absence of the protein and then normalized to 100% translational activity. As shown in Fig. 2, we observed that MCL1 was able to effectively reduce the yield of luciferase and a presumed IC50 value for MCL1 was 1.9 nM. These results indicate that MCL1 could possibly be classified as a type-2 RIP.
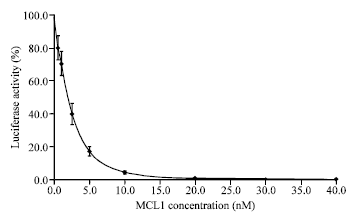 | |
| Fig. 2: | Ribosome-inactivating activity of MCL1. Ribosome-inactivating activity was determined by measuring the degree of inhibition of luciferase translation in a rabbit reticulocyte lysate system. The luciferase activity was normalized to 100% in the absence of MCL1. Results are given (Mean±SE) for 3 experiments consisting of duplicate samples |
There were nine potential N-linked glycosylation sites (Asn-X-Ser/Thr) in the deduced amino acid sequence (Fig. 1). Five QxW repeats were observed in the B-chain of MCL1, which may be involved in its tolerance to accommodate greatly differing amino acids at most positions in the structure (Hazes, 1996). To check whether MCL1 has introns, genomic DNA from M. charantia was amplified by PCR using oligonucleotide primers corresponding to the N- and C-terminal sequences of the primary translation product of MCL1 (5'-ATGAGAATGAGAGTGTTA-3' and 5'-ATAGAACACCGTCCATTG-3'). Analysis of PCR products confirmed that the 1.7 kbp amplified fragment was the same size as a fragment amplified by using the cDNA template. Moreover, the sequence of this amplicon was identical to the cDNA encoding MCL1. These results suggest that no intron exists in the coding region of MCL1, a result that is similar to many other reported type-2 RIPs (Halling et al., 1985; Wood et al., 1991; Kaku et al., 1996; Peumans et al., 1998).
Sequence comparisons with other lectins: A BLASTP search for MCL1 in the protein sequence database found significant degrees of identity to the following type-2 RIPs: 40 and 39% for Sambucus nigra SNAI' and SNAV (Van Damme et al., 1996, 1997), respectively; Abrus precatorius Abrin-c (36%, Wood et al., 1991) and Ricinus communis Ricin (34%, Halling et al., 1985). The alignment of some representatives of these homologous sequences is shown in Fig. 3. These results show that MCL1 was different from other reported lectins. Although Glu177 and Arg180 play an important role in the rRNA-N-glycosidase activity of ricin A-chain (Katzin et al., 1991) and were highly conserved in this study, the Arg186 residue was replaced by Lys186 in MCL1. Moreover, Katzin et al. (1991) also reported several polar residues involved in the catalysis. Tyrosines 80 and 123 in ricin were replaced by nonpolar Gly95 and Phe134, respectively. The phylogenetic trees that were built based on the amino acid sequences of A- and B-chains of type-2 RIPs from these different species indicate that the A-chain of MCL1 is comparatively more distantly related to these species, whereas the B-chain is evolutionary closer to that of Sambucus lectins (Fig. 4). MCL1 seemed to be phylogenetically distant from the other type-2 RIPs, because evolutionary relationships between A- and B-chain are similar in this class of RIPs, except for MCL1.
Tissue distribution of MCL1 mRNA: RT-PCR and real-time PCR were performed to determine the tissue-specific expression of MCL1 mRNA. The RT-PCR products were cloned and sequenced to confirm the specificity of RT-PCR amplification and identity of the expected sequences. Xu et al. (2007) reported that the RIP coding gene cloned and sequenced by Lee-Huang et al. (1995) from M. charantia was constitutively expressed in seeds, flowers, roots and stems, whereas the MCL1 transcript was detected only in the embryos (Fig. 5).
 | |
| Fig. 3: | Alignment of amino acid sequences of M. charantia MCL1 and other homologous type-2 RIPs. The alignment was performed using the ClustalW program. Dashes indicate gaps introduced during the alignment process. Asterisks indicate identity and single and double dots indicate semi-conservative and conservative replacements, respectively. Numbering of the amino acids starts at the N-termini of the proteins. Cysteine residues potentially involved in disulfide-bridges are indicated by bold letters. The boxed regions are described in the text. MCL1, Momordica charantia (this study); SNAV, Sambucus nigra (U41299); Ricin, Ricinus communis (X03179); AbrinC, Abrus precatorius (X55667) |
Moreover, similar results were obtained using real-time PCR (data not shown). The hemagglutinating activity was detected in embryos and cotyledons, but was not detected in leaves or stems. The results of this study show that MCL1 gene expression occurs only during embryogenesis and the product accumulates in embryos and cotyledons. The translation product of MCL1 may play an important role in the protection of seeds against insects and microorganisms.
Expression of M. charantia MCL1 in tobacco BY-2 cells: In order to prove the identity of MCL1 as a lectin gene, the expression of MCL1 cDNA in tobacco BY-2 suspension cells was studied using crude extracts of the cells. The supernatant fluid of the disrupted cells showed higher hemagglutinating activity toward human type-O(H) erythrocyte than the other tested erythrocytes, whereas the control transformant without the MCL1 gene showed no detectable activity. Moreover, the carbohydrate-binding specificity of the lectin was studied by carrying out using saccharide inhibition assays. The following saccharides did not inhibit hemagglutination at concentrations up to 100 mM: L-fucose, D-mannnose, D-fructose and sucrose.
 | |
| Fig. 4: | Phylogenetic relationships between M. charantia MCL1 and homologous type-2 RIPs. The phylogenetic trees were built up based on the amino acid sequences of A- and B-chains of type-2 RIPs from different species. The values represent the percentage of 1,000 bootstrap replications. MCL1, Momordica charantia (this study); LPRSN1, SNAI and SNAV, Sambucus nigra (U58358, U27122, U41299); Ebulin1, Sambucus ebulus (AJ400822); PMRIPt, Polygonatum multiflorum (AF213984); Viscumin1, Viscum album (AY377890); Ricin, Ricinus communis (X03179); AbrinC, Abrus precatorius (X55667); Cinnamomin, Cinnamomum camphora (AY039801). Panel A, phylogenetic tree of A-chain; panel B, phylogenetic tree of B-chain |
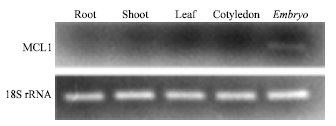 | |
| Fig. 5: | Tissue distributions of MCL1 mRNA expression in various tissues analyzed by RT-PCR. 18S rRNA was used as the internal control. Total RNA extracted from individual tissues was reverse transcribed and used for PCR analysis of MCL1 and 18S rRNA mRNA expression |
Human blood type-H antigen trisaccharide showed the strongest inhibition of all tested reagents. These results suggest that MCL1 was encoding a galactose-specific anti-H lectin and the binding specificity resembles the purified lectin (Toyama et al., 2008).
In conclusion, we have cloned an MCL1, coding anti-H lectin, from the seeds of M. charantia and successfully expressed this type-2 RIP protein in tobacco BY-2 cells for the first time. The transgenic tobacco cells and the expressed MCL1 are expected to be useful in blood typing (Toyama et al., 2008), cell activator (Huang et al., 2008) and other applications.
ACKNOWLEDGMENTS
We wish to acknowledge Dr. Yoshihiko Tani and Ms. Junko Takahashi at the Osaka Red Cross Blood Center, Japan for useful discussions and technical advice. Tobacco BY-2 cells were provided by RIKEN BRC, which is participating in the National Bio-Resource Project of the Ministry of Education, Culture, Sports, Science and Technology, Japan. This research was supported by the Japan Science and Technology Agency (JST) Practical Application Research program.
REFERENCES
- Barbieri, L., M. Zamboni, E. Lorenzoni, L. Montanaro, S. Sperti and F. Stirpe, 1980. Inhibition of protein synthesis in vitro by proteins from the seeds of Momordica charantia (bitter pear melon). Biochem. J., 186: 443-452.
PubMed - Goldstein, I.J., R.C. Hughes, M. Monsigny, T. Osawa and N. Sharon, 1980. What should be called a lectin? Nature, 285: 66-66.
CrossRef - Gondo, T., S.I. Tsuruta, R. Akashi, O. Kawamura and F. Hoffmann, 2005. Green, herbicide-resistant plants by particle inflow gun-mediated gene transfer to diploid bahiagrass (Paspalum notatum). J. Plant Physiol., 162: 1367-1375.
CrossRefDirect Link - Halling, K.C., A.C. Halling, E.E. Murray, B.F. Ladin, L.L. Houston and R.F. Weaver, 1985. Genomic cloning and characterization of a ricin gene from Ricinus communis. Nucleic Acids Res., 13: 8019-8033.
CrossRefDirect Link - Kaku, H., Y. Tanaka, K. Tazaki, E. Minami, H. Mizuno and N. Shibuya, 1996. Sialylated oligosaccharide-specific plant lectin from Japanese elderberry (Sambucus sieboldiana) bark tissue has a homologous structure to type II ribosome-inactivating proteins, ricin and abrin. J. Biol. Chem., 271: 1480-1485.
- Kumar, S., K. Tamura and M. Nei, 2004. MEGA3: Integrated software for molecular evolutionary genetics analysis and sequence alignment. Brief. Bioinform., 5: 150-163.
CrossRefPubMedDirect Link - Lee-Huang, S., P.L. Huang, H.C. Chen, P.L. Huang, A. Bourinbaiar, H.I. Huang and H.F. Kung, 1995. Anti-HIV and anti-tumor activities of recombinant MAP30 from bitter melon. Gene, 161: 151-156.
PubMed - Melo, V.M.M., I.M. Vasconcelos, V.M. Gomes, M.D. Cunha, A.A. Soares and J.T.A. Oliveira, 2005. A cotyledonary agglutinin from Luetzelburgia auriculata inhibits the fungal growth of Colletotrichum lindemuthianum, Fusarium solani and Aspergillus niger and impairs glucose-stimulated acidification of the incubation medium by Saccharomyces cerevisiae cells. Plant Sci., 169: 629-639.
CrossRef - Toyama, J., H. Tanaka, A. Horie, T. Uchiyama and R. Akashi, 2008. Purification and characterization of Anti-H lectin from the seed of Momordica charantia and the inter-specific differences of hemagglutinating activity in Cucurbitaceae. Asian J. Plant Sci., 7: 647-653.
CrossRefDirect Link - Wang, H. and T.B. Ng, 1998. Ribosome inactivating protein and lectin from bitter melon (Momordica charantia) seeds: Sequence comparison with related proteins. Biochem. Biophys. Res. Commun., 253: 143-146.
PubMed - Wong, J.H., C.C. Wong and T.B. Ng, 2006. Purification and characterization of a galactose-specific lectin with mitogenic activity from pinto beans. Biochim. Biophys. Acta, 1760: 808-813.
CrossRefDirect Link