
Kayode A. Sanni
Africa Rice Center (WARDA), 01 BP 2031, Cotonou, Benin Republic
O.J. Ariyo
Department of Plant Breeding and Seed Technology, University of Agriculture, Abeokuta, P.M.B. 2240, Abeokuta, Nigeria
D.K. Ojo
Department of Plant Breeding and Seed Technology, University of Agriculture, Abeokuta, P.M.B. 2240, Abeokuta, Nigeria
G. Gregorio
International Rice Research Institute, Manila, Philippines
E.A. Somado
Africa Rice Center (WARDA), 01 BP 2031, Cotonou, Benin Republic
I. Sanchez
Africa Rice Center (WARDA), 01 BP 2031, Cotonou, Benin Republic
M. Sie
Africa Rice Center (WARDA), 01 BP 2031, Cotonou, Benin Republic
K. Futakuchi
Africa Rice Center (WARDA), 01 BP 2031, Cotonou, Benin Republic
S.A. Ogunbayo
Africa Rice Center (WARDA), 01 BP 2031, Cotonou, Benin Republic
R.G. Guei
Food and Agricultural Organization, Viale delle Terme de Caracalla, Rome 00100, Italy
M.C.S. Wopereis
Africa Rice Center (WARDA), 01 BP 2031, Cotonou, Benin Republic
Asian Journal of Plant Sciences
Year: 2009 | Volume: 8 | Issue: 1 | Page No.: 48-53







ABSTRACT
Genotype by Environment Interaction (GEI) is a major complications in plant breeding. We used Additive Main Effects and Multiplicative Interaction (AMMI) to evaluate the effects of GEI in NERICA rice genotype and their adaptation in two years at three locations; Ibadan-7°30’ N, 3°58’ E, 210 m.a.s.l. (Nigeria), Cotonou-6°24’ N, 2°19’ E, 15.5 m.a.s.l. and Deve-6°48’ N, 1°47’ E, 72 m.a.s.l. (Benin Republic). Twenty two rice genotypes were grown in 2005 and 2006 under upland condition, using randomized complete block design with three replications. Main effects due to environments (E), genotypes (G) and GxE interaction (GEI) were significant (p<0.01), with the highest variation of 43.1% accounted for by environmental effects. The first four Interaction Principal Component Axes (IPCA1, 2, 3 and 4) were significant (p<0.01) and cumulatively contributed 98.5% of the total GEI. AMMI biplot accounted for 91.4% of the total sum of squares. The stability study indicated that NERICAs 3, 10, 11 and 18 could be considered stable in any of the environments, due to their low interactions. However, NERICA 11 was the most promising of the genotypes, with high yield (5.15 t ha-1) and a broad environmental adaptation.
PDF Abstract XML References Citation
How to cite this article
Kayode A. Sanni, O.J. Ariyo, D.K. Ojo, G. Gregorio, E.A. Somado, I. Sanchez, M. Sie, K. Futakuchi, S.A. Ogunbayo, R.G. Guei and M.C.S. Wopereis, 2009. Additive Main Effects and Multiplicative Interactions Analysis of Grain Yield Performances in Rice Genotypes Across Environments. Asian Journal of Plant Sciences, 8: 48-53.
DOI: 10.3923/ajps.2009.48.53
URL: https://scialert.net/abstract/?doi=ajps.2009.48.53
DOI: 10.3923/ajps.2009.48.53
URL: https://scialert.net/abstract/?doi=ajps.2009.48.53
INTRODUCTION
Rice is grown and consumed in about 39 countries of Africa. Fuelled by changing lifestyle, population growth and consumer preference, the demand for rice in West and Central Africa (WCA), is growing at the rate of 6% per annum, which is fasterthan anywhere else in the world, while production increase at the rate of 4% per annum (WARDA, 2004). Africa consumes a total of 11.6 million tonnes of milled rice per year (FAO, 1996), of which 3.3 million tonnes (33.6%) is imported. Among the 39 African countries that produce and consume rice, only ten have attained any appreciable levels of rice self-sufficiency (75-99.9%), while the remaining 29 are heavy importers with self-sufficiency levels ranging between 0 and 62.8% (Oteng and Sant’Anna, 1999). There is no doubt that rice production in Africa must increase if the high demand in Africa will be met.
One of the main objectives of rice improvement in Sub-Saharan Africa (SSA) is to improve yield and increase local production. However, its production in Sub-Saharan Africa (SSA) has been limited by the typical African stresses (biotic and abiotic). Oryza consists of two cultivated species Oryza sativa which originates from Asia and O. glaberrima which is indigenous to Africa. O. glaberrima which has been discovered and cultivated in Africa over 3500 years ago (Chang, 1976) is well adapted to the various adverse conditions of Africa, such as the blast, rice yellow mottle virus, rice gall midge, insects, nematodes, acidity, iron toxicity, salinity and drought, but with poor yield. Although, the Asian rice (O. sativa) which was introduced to Africa about 2000 years ago has a better yield but it is not sufficiently adapted to the typical African stresses.
Research work at the Africa Rice Centre (WARDA) has led to a major success with the development of improved upland rice varieties from interspecific cross between O. sativa and O. glaberrima to produce what is now referred to as the New Rice for Africa (NERICA). NERICA has great potential to benefit African farmers because of its high-yielding ability under typical upland conditions in this region (Ikeda et al., 2007). Eighteen of these NERICA varieties have been named by WARDA and are being widely disseminated across the region by WARDA and its partners.
The adoption of these named NERICA varieties is increasingly spreading in the upland rice-based systems, which covers the most extensive rice ecosystem in SSA. As a result of this, it is therefore important to assess the adaptation and yield stability of these promising upland genotypes of rice across environments.
Several statistical methods have been developed for the analysis of Genotype by Environment Interactions (GEI) and phenotypic stability (Hill, 1975; Lin et al., 1986; Wescott, 1986; Crossa, 1990; Flores et al., 1998). Regression technique has been widely used (Eberhart and Russell, 1966; Perkins and Jinks, 1968) due to its simplicity and the fact that its information on adaptive response is easily applicable to locations (Annicchiarico, 1997). The Principal Component Analysis (PCA) method that shows the mean squares of the principal components axes (Gauch and Furnas, 1991) has also been used. Zobel et al. (1988) compared the traditional statistical analysis such as Analysis of Variance (ANOVA), Principal Component Analysis (PCA) and Linear Regression with AMMI analyses, and showed that the traditional analyses were not always effective in analyzing the multi-environment trial data structure. The ANOVA is an additive model that describes main effects effectively and determines if GE interaction is a significant source of variation, but it does not provide insight into the patterns of genotypes or environments that give rise to the interaction. The PCA is a multiplicative model that contains no sources of variation for additive G or E main effects and does not analyze the interactions effectively. The linear regression method uses environmental means, which are frequently a poor estimate of environments, such that the fitted lines in most cases account for a small fraction of the total GE and could be misleading (Byth et al., 1976; Zobel et al., 1988; Ariyo, 1999).
Additive main effects and multiplicative interaction (AMMI) has been proved to be a suitable method for depicting adaptive responses (Gauch, 1993; Annicchiarico, 1997; Gauch and Zobel, 1989; Ariyo, 1999). AMMI analysis has been reported to have significantly improved the probability of successful selection (Gauch and Zobel, 1989) and has been used to analyse GxE interaction with greater precision in many crops (Bradu, 1984; Gauch 1990; Crossa et al., 1991; Ariyo, 1999). The model combines the conventional analysis of variance for genotype and environment main effects with principal components analysis to decompose the GEI into several Interaction Principal Component Axes (IPCA). With the biplot facility from AMMI analysis, both genotypes and environments are plotted together on the same scatter plot and inferences about their interaction can be made.
This study reports the use of AMMI model to analyse yield data of twenty two genotypes of rice from two years evaluation in three locations. The objective was to quantify genotypexenvironment interaction for grain yield and the stability of NERICA rice genotypes in different environments.
MATERIALS AND METHODS
Plant materials: Twenty two rice genotypes comprising of the 18 named interspecific NERICA rice varieties (Oryza sativaxO. glaberrima) and their parents O. glaberrima steud (CG 14) and O. sativa L. (WAB56-104, WAB56-50 and WAB181-18) were used in this study (Table 1). Apart from CG 14 which is a landrace from Senegal, all the genotypes originated from WARDA, Bouaké, Côte d’Ivoire. The seeds were obtained from the rice germplasm collection of the Genetic Resources Unit at the Africa Rice Center (WARDA).
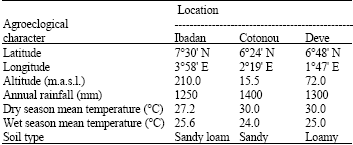
Field trials: The experiment was carried out during the rainy seasons (June to October) of 2005 and 2006 under upland condition at three locations in two West African countries-Nigeria (Ibadan) and Benin (Cotonou and Deve). The geographical coordinates and agroclimatic characteristics of the locations are shown in Table 2. At each location, the genotypes were grown through direct seeding, by dibbling in randomized complete block design with three replications. The size of each plot was 1x5 m made up of 6 rows, with a spacing of 20 cm apart. The inter-plot spacing was 50 cm. Fertilizer application at each site consisted of NPK (10-18-18) used as basal application at the rate of 100 kg ha-1 during land preparation. Urea (N: 46%) was applied at the rate of 65 kg ha-1 as top-dressing, first at tillering and second at panicle development (booting) stage. The plots were hand-weeded regularly to minimize weed infestation. Bird damage was controlled using bird scares.
Statistical analysis: Data on grain yield for the 22 genotypes of rice in 6 year-location environments were subjected to AMMI analysis. To implement the AMMI, the data was input into the microcomputer software program MATMODEL Version 2.2. The AMMI model is:
Yger = μ+αg+βe+Σλnγgnηen+�?ge+�μger |
| Table 1: | List of the studied genotypes |
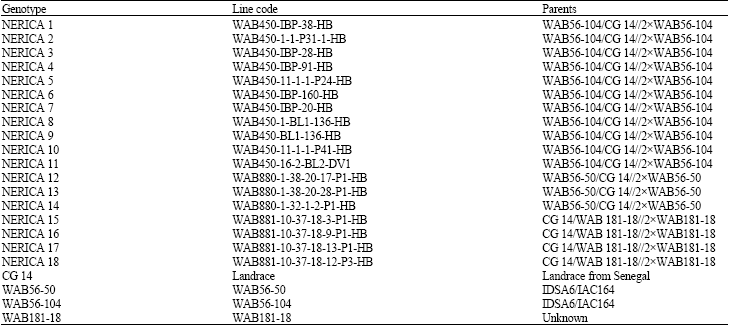 | |
IBP: Interbreeding population, HB: High input homogeneous bulk, P: Panicle selection, BL: Blast, DV: Drought at vegetative phase | |
| Table 2: | Description of the three experimental locations |
 | |
where, Yger is the yield of genotype g in environment e for replicate r, μ is the grand mean, αg is genotype mean deviation (mean minus the grand mean), βe is the environment mean deviations, N is the number of PCA axes retained in the model, λn is the singular value of PCA axis n, γgn is the genotype eigenvector values for PCA axis n, ηen is the environment eigenvector values for PCA axis n, I?ge is the AMMI residuals and εger is the residual error.
AMMI uses ordinary ANOVA to analyse the main effects (additive part) and Principal Component Analysis (PCA) to analyse the non additive residual left over by the ANOVA (Gauch, 1993). In the analysis, each combination of a single location and year was considered as an environment.
The interaction is the genotype PCA score multiplied by that of the environment. When a cultivar and an environment have the same sign on their respective first PCA axis, their interaction is positive; if different, their interaction is negative. An AMMI plot is a graph where aspects of both genotypes and environments are plotted on the same axis so that interrelationship can be visualised. It provides a pictorial view of the transformed GxE interaction (Kempton, 1984) for easy interpretation. In a biplot where, PCA 1 score is on the vertical axis and the mean yield on the horizontal, genotypes that appear almost on a perpendicular line had similar means and those that fall almost on a horizontal line had similar interaction patterns. Similarly, environment that occurred almost on a perpendicular line had similar means and those on horizontal lines had similar interaction patterns. Genotypes or environments with large PCA 1 scores, either positive or negative had large interactions, whereas genotypes with PCA 1 score of zero or nearly zero had smaller interaction (Crossa et al., 1990). The biplot of the first two IPCA axes demonstrates the relative magnitude of the GEI for specific genotypes and environments. The further away from the axes center a genotype or environment is, the larger the GEI.
RESULTS AND DISCUSSION
Genotype (G) which accounted for 41.6% of the total Sum of Square (SS), environmental effects (E) which explained 43.1% and the GxE Interaction (GEI) effects which captured 15.3%, were all significant (p = 0.01), indicating that all sources are important in the analysis (Table 3). The result showed that the environment main effect (E) was the most important source of variation, due to its large contribution to the total sum of squares for yield (Kaya et al., 2002). Variation due to G was larger than that due to GEI, but GEI was significant p = 0.01, meaning that differences among genotypes vary across environments (Admassu et al., 2008).
The presence of GEI was clearly demonstrated by the AMMI model, when the interaction was partitioned among the first four Interaction Principal Component Axis (IPCA) as they were significant p = 0.01 in a postdictive assessment.
| Table 3: | AMMI analysis of variance for rice grain yield |
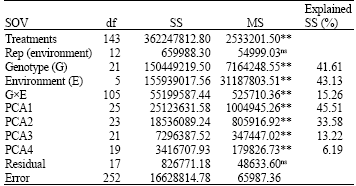 | |
**Significant at 1% probability level, ns: Not significant | |
The IPCA1 explained 45.51% of the interaction sum of square in 25% of the interaction degree of freedom (df). Similarly, the second, third and fourth principal component axis (PCA 2-4) explained a further 33.58, 13.2 and 6.19% of the GEI sum of square, respectively. They cumulatively captured 98.50% of the total GEI SS, using 88 df. This implied that the interaction of the 22 genotypes of rice with six environments was predicted by the first four principal components of genotypes and environments, which is in agreement with the recommendation of Sivapalan et al. (2000). This is also in concordance with the results of Van Oosterom et al. (1993), where as much as the first five IPCAs were significant. However, this contradicted the findings of Gauch and Zobel (1996), which recommended that the most accurate model for AMMI, can be predicted using the first two IPCAs. These results indicate that the number of the terms to be included in an AMMI model cannot be specified a prior without first trying AMMI predictive assessment (Kaya et al., 2002).
The AMMI analysis provided a biplot (Fig. 1) of main effects and the first principal component scores of interactions (IPCA1) of both genotypes and environments. The differences among genotypes in terms of direction and magnitude along the X-axis (yield) and Y axis (IPCA 1 scores) are important. In the biplot display, genotypes or environments that appear almost on a perpendicular line of the graph had similar mean yields and those that fall almost on a horizontal line had similar interaction (Crossa et al., 1990). Thus the relative variability due to environments was greater than that due to genotype differences. Genotypes or environments on the right side of the midpoint of the perpendicular line have higher yields than those on the left side. Consequently, NERICA11, 10, 17, 16 and 9 were generally high yielding (5.15, 4.71, 4.55, 4.45 and 4.36 t ha-1, respectively) with NERICA11 being the overall best with yield of 5.15 t ha-1 (Fig. 1). In contrast, CG 14, WAB56-50 and WAB56-104 were generally low yielding genotypes.
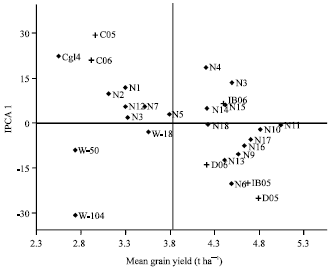 | |
| Fig. 1: | Biplot of grain yield of 22 rice genotypes and environments AMMI plot for rice grain yield trials with 22 genotypes grown in six environments. IPC1: First-interaction principal component, C: Cotonou, D: Deve; IB: Ibadan, N: NERICA, W-18: WAB181-18; W-50: WAB56-50, W-104: WAB56-104 |
The Deve and Ibadan locations which were always on the right hand side of the midpoint of the main effect axis, seemed to be favourable environments, with Deve as the most favourable, while Cotonou is the less favourable environment.
Genotypes or environments with large negative or positive IPCA1 scores have high interactions, while those with IPCA1 scores near zero (close to the horizontal line) have little interaction across environments and vice versa for environments (Crossa et al., 1991; Egesi and Asiedu, 2002) and are considered more stable than those further away from the line. In the biplot, NERICA11, 10 and 18 fell almost on a horizontal line near the zero point on IPCA1, but were vertically distant apart. This implies that NERICA 11 and 10 showed high and stable yield; yield of NARICA 18 is also stable against environmental changes but its potential is lower than NERICA 11 and 10.
Since, the IPCA 2 scores was also important (33.58% of GxE SS) in explaining GEI, the biplot of the first two IPCAs was also used to demonstrate the relative magnitude of the GEI for specific genotypes and environments (Fig. 2). The IPCA scores of genotypes in the AMMI analysis is an indication of stability or adaptation over environments (Gauch and Zobel, 1996). The greater the IPCA scores, the more specifically adapted is a genotype to certain environments (Admassu et al., 2008). The more the IPCA scores approximate to zero, the more stable or adapted the genotype is over all the environments sampled. A biplot of the first two IPCA show that the best adapted genotype to most environments was NERICA11 with the IPCA scores close to zero, indicating its stability over the entire environments. WAB56-104 and WAB181-18 were well adapted to the high yielding environments of Deve- 2005 and 2006, while CG 14 was well adapted in the low yielding environments of Cotonou 2005 and 2006. NERICA15 and 14 performed best at Ibadan in 2006 while the best performing variety at Ibadan in 2005 was NERICA6.
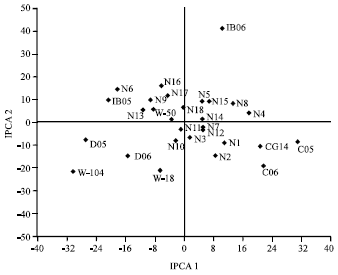 | |
| Fig. 2: | Biplot of the second interaction principal component axis (IPCA 2) against the first interaction principal component axis (IPCA 1) scores for grain yield of 22 rice genotypes in six environments. C = Cotonou, D = Deve, IB = Ibadan, N = NERICA, W-18 = WAB181-18, W-50 = WAB56-50, W-104 = WAB56-104 |
Considering the environments tested in this study, no single location had IPCA 1 scores close to zero line. Also the IPCA 2 scores for environments were far from zero. This indicates that all the environments had potential for large GEI.
CONCLUSION
The AMMI statistical model has shown that the largest proportion of the total variation in grain yield was attributed to environments in this study. NERICA 11 and 10 had the highest yield and were hardly affected by the GEI effects as a result of which they will perform well across a wide range of environments. The yield of NARICA 18 was also stable against environmental changes but its potential was lower than NERICA 11 and 10.
REFERENCES
- Admassu, S., M. Nigussie and H. Zelleke, 2008. Genotype-environment interaction and stability analysis for grain yield of maize (Zea mays L.) in Ethiopia. Asian J. Plant Sci., 7: 163-169.
CrossRefDirect Link - Annicchiarico, P., 1997. Joint regression vs. AMMI analysis of genotype-environment interactions for cereals in Italy. Euphytica, 94: 53-62.
CrossRef - Chang, T.T., 1976. The origin, evolution, cultivation, dissemination and diversification of Asian and African rices. Euphytica, 25: 425-441.
CrossRef - Crossa, J., H.G. Gauch and R.W. Zobel, 1990. Additive main effects and multiplicative interaction analysis of two international maize cultivar trials. Crop Sci., 30: 493-500.
CrossRefDirect Link - Crossa, J., P.N. Fox, W.H. Pfeiffer, S. Rajaram and H.G. Gauch Jr., 1991. AMMI adjustment for statistical analysis of an international wheat yield trial. Theoret. Applied Genet., 81: 27-37.
CrossRefDirect Link - Egesi, C.N. and R. Asiedu, 2002. Analysis of yam yields using the additive main effects and multiplicative interaction (AMMI) model. Afr. Crop Sci. J., 10: 195-201.
Direct Link - Oteng, J.W. and R. Sant'Anna, 1999. Rice production in Africa: Current situation and issues. Int. Rice Com. Newsl., 48: 41-51.
Direct Link - Flores, F., M.T. Moreno and J.I. Cubero, 1998. A comparison of univariate and multivariate methods to analyze GxE interaction. Field Crops Res., 56: 271-286.
CrossRefDirect Link - Byth, D.E., R.L. Eisemann and I.H. de Lacy, 1976. Two-way pattern analysis of a large data set to evaluate genotypic adaptation. Heredity, 37: 215-230.
CrossRefDirect Link - Eberhart, S.A. and W.A. Russell, 1966. Stability parameters for comparing varieties. Crop Sci., 6: 36-40.
CrossRefDirect Link - Hill, J., 1975. Genotype-environment interactions-a challenge for plant breeding. J. Agric. Sci. Camb., 85: 477-493.
CrossRef - Gauch, H.G. Jr., 1990. Full and reduced models for yield trials. Theor. Applied Genet., 80: 153-160.
CrossRef - Ikeda, R., Y. Sokei and I. Akintayo, 2007. Reliable multiplication of seed for NERICA varieties of rice, Oryza sativa L. Genet. Resour. Crop Evol., 54: 1637-1644.
CrossRefDirect Link - Kaya, Y., C. Palta and S. Taner, 2002. Additive main effects and multiplicative interactions analysis of yield performance in bread wheat genotypes a cross environments. Turk. J. Agric., 26: 275-279.
Direct Link - Kempton, R.A., 1984. The use of biplots in interpreting variety by environment interactions. J. Agric. Sci., 103: 123-135.
CrossRefDirect Link - Gauch, H.G. and R.E. Furnas, 1991. Statistical analysis of yield trials with MATMODEL. Agron. J., 83: 916-920.
Direct Link - Lin, C.S., M.R. Binns and L.P. Lefkovitch, 1986. Stability analysis: Where do we stand? Crop Sci., 26: 894-900.
CrossRefDirect Link - Sivapalan, S., L.O. Brien, G. Ortiz-Ferrara, G.L. Hollamby, I. Barclay and P.J. Martin, 2000. An adaptation analysis of Australian and CIMMYT/ICARDA wheat germplasm in Australian production environments. Aust. J. Agric. Res., 51: 903-915.
CrossRefDirect Link - Van Oosterom, E.J., D. Kleijn, S. Ceccarelli and M.M. Nachit, 1993. Genotype-by-environment interactions of barley in the mediterranean region. Crop Sci., 33: 669-674.
CrossRefDirect Link - Wescott, B., 1986. Some methods of analyzing genotype-environment interaction. Heredity, 56: 243-253.
CrossRefDirect Link - Perkins, J.M. and J.L. Jinks, 1968. Environmental and genotype-environmental components of variability III. Multiple lines and crosses. Heredity, 23: 339-356.
CrossRef - Gauch Jr., H.G. and R.W. Zobel, 1989. Accuracy and selection success in yield trials analysis. Theor. Applied Genet., 77: 443-481.
CrossRefDirect Link
Ronoubigouwa Michael Reply
Thank you for this great and very informative job. It would be useful for the dissemination of NERICAs and their growth in many different environments and ecosystems.
imam muhammad bello Reply
the paper is good enough for me to have a glance of it. i will need the full paper to go through it.