
P. Songsri
Department of Plant Science and Agricultural Resources, Faculty of Agriculture, Khon Kaen University, Muang, Khon Kaen, 40002, Thailand
N. Vorasoot
Department of Plant Science and Agricultural Resources, Faculty of Agriculture, Khon Kaen University, Muang, Khon Kaen, 40002, Thailand
S. Jogloy
Department of Plant Science and Agricultural Resources, Faculty of Agriculture, Khon Kaen University, Muang, Khon Kaen, 40002, Thailand
T. Kesmala
Department of Plant Science and Agricultural Resources, Faculty of Agriculture, Khon Kaen University, Muang, Khon Kaen, 40002, Thailand
C. Akkasaeng
Department of Plant Science and Agricultural Resources, Faculty of Agriculture, Khon Kaen University, Muang, Khon Kaen, 40002, Thailand
A. Patanothai
Department of Plant Science and Agricultural Resources, Faculty of Agriculture, Khon Kaen University, Muang, Khon Kaen, 40002, Thailand
C.C. Holbrook
USDA-ARS, Coastal Plain Experiment Station, P.O. Box 748, Tifton, 31793, GA, USA
Asian Journal of Plant Sciences
Year: 2009 | Volume: 8 | Issue: 7 | Page No.: 465-473







ABSTRACT
The aim of this study was to evaluate the responses to difference in available soil water levels for yield and reproductive characters of peanut genotypes and relate these responses to pod yield under drought conditions. Eleven peanut genotypes were tested under three soil moisture levels (Field Capacity (FC), 2/3 available soil water (AW) and 1/3AW). Data were recorded for total number of flowers, pegs (reproductive sinks; RSs), immature pods and mature pods per plant, number of seeds per pod, 100-seed weight and pod yield at harvest. Drought at mild and severe levels significantly reduced Harvest Index (HI), yield components and reproductive developmental characters and the reductions were most substantial for pod yield with increasing moisture stress. High pod yield under drought conditions in ICGV 98300 was caused by high potential yield and low yield reduction, whereas high pod yield in ICGV 98324 was due to low pod yield reduction. High HI and numbers of mature pods are advantageous and necessary for high yield under drought conditions. High conversion of RSs to total pods and conversion of flowers to mature pods were the most important factors contributing to high pod yield under mild drought (2/3 AW) and severe drought (1/3 AW), respectively. Tifton 8 showed the lowest pod yield and poor seed filling under well-watered and droughts conditions.
PDF Abstract XML References Citation
How to cite this article
P. Songsri, N. Vorasoot, S. Jogloy, T. Kesmala, C. Akkasaeng, A. Patanothai and C.C. Holbrook, 2009. Evaluation of Yield and Reproductive Efficiency in Peanut (Arachis hypogaea L.) under Different Available Soil Water. Asian Journal of Plant Sciences, 8: 465-473.
DOI: 10.3923/ajps.2009.465.473
URL: https://scialert.net/abstract/?doi=ajps.2009.465.473
DOI: 10.3923/ajps.2009.465.473
URL: https://scialert.net/abstract/?doi=ajps.2009.465.473
INTRODUCTION
More than 80% of world’s peanut production comes from rainfed agriculture (Wright and Nageswara Rao, 1994). Erratic and insufficient rainfall is a major constraint of peanut production in rain-fed environments and water is increasingly becoming a scarce resource even in irrigated agriculture because of increasing demands for urban and industrial consumptions. Drought stress during the stages of pod and seed formation has shown to reduce pod yield (Nageswara Rao et al., 1989; Reddy et al., 2003; Vorasoot et al., 2003) and increase the likelihood of aflatoxin contamination (Rachaputi et al., 2002; Holbrook and Stalker, 2003).
The relationships between pod yield and reproductive characters may be altered by drought. So far, the response of genotype to drought in relation to reproductive characters and pod yield are not well understood. Previous studies indicated that the occurrence of drought during the vegetative phase has only small effect on growth and yield of peanut (Nageswara Rao et al., 1988; Nautiyal et al., 1999). However, drought during the flowering and pod formation phases is severely detrimental to the yield of peanut (Nautiyal et al., 1999; Wright and Nageswara Rao, 1994) as it lowered (Awal and Ikeda, 2002). Under water deficit conditions, pod yield was affected by decreasing pod growth and development (Reddy et al., 2003; Chapman et al., 1993) and drought also decreased number of mature pods and pod yield (Nautiyal et al., 1999).
Pod yield can be considered as the sequential processes of flower production, peg initiation, conversion of peg to pods and pod filling. To the best of our knowledge, the reproductive efficiency such as the conversion of flowers, pegs and mature pods to pod yield under drought have not been investigated thoroughly.
Therefore, the objectives of this study were to evaluate the responses of peanut genotypes to differences in available soil water levels for yield, yield components and reproductive development characters and to relate these responses to pod yield under drought conditions. Information of the study would provide a better understanding on how genotypes could achieve high yield under drought and will have important implications for breeding for drought resistance in peanut.
MATERIALS AND METHODS
Pot experiment was conducted under greenhouse conditions at the Field Crop Research Station of Khon Kaen University located in Khon Kaen province (latitude 16°28' N, longitude 102° 48' E, 200 m above sea level) during December 2002 to May 2003 (GH1) and repeated during June to November 2003 (GH2).
Plant materials and experimental procedures: Eleven peanut genotypes were used in this study. Eight (ICGV 98300, ICGV 98303, ICGV 98305, ICGV 98308, ICGV 98324, ICGV 98330, ICGV 98348 and ICGV 98353) are elite drought resistant lines obtained from ICRISAT, one (Tifton-8) is a Virginia-type drought resistant line (Coffelt et al., 1985) received from the United State Department of Agriculture (USDA) and two (KK 60-3 and Tainan 9) are released cultivars commonly grown in Thailand. The lines from ICRISAT were identified as drought resistant because they produced high total biomass and pod yield in screening tests under drought conditions (Nageswara Rao et al., 1992; Nigam et al., 2003, 2005). KK 60-3 is a Virginia-type peanut cultivar sensitive to drought for pod yield, while Tainan 9 is a Spanish-type peanut cultivar having low dry matter production (Vorasoot et al., 2003, 2004).
A 3 x 11 factorial combination in a Randomized Complete Block Design (RCBD) with six replications was used for both experiments. Three soil moisture levels (Field Capacity (FC), 2/3 available soil water (2/3 AW) and 1/3 available soil water (1/3 AW)) were assigned as factor A and 11 peanut genotypes as factor B.
The soil on the experimental site pertains to the Yasothon series (Yt; fine-loamy, siliceous, isohypo-thermic, Oxic Paleustults). The proportions of sand, silt and clay in the soil were 56.84, 24.79 and 18.37%, respectively. A sandy loam soil with pH 5.20, 0.196% organic matter and 0.0093% total N. Available P was 4.88 ppm (Bray II method) and extractable K and Ca were 49.55 and 444.94 ppm, respectively.
A pot size of 25x70 cm was used for growing peanut. Each pot was filled with 42 kg dry soil to 10 cm from the top for creating the uniform bulk density. Each treatment consisted of two pots in a replicate. Seeds were treated with captan (3a, 4, 7, 7a-tetrahydro-2-((trichloromethyl)thio)-1H-isoindole-1, 3(2H)-dione) at the rate of 5 g kg-1 seed before planting and seeds of the two Virginia-type peanut genotypes (KK 60-3 and Tifton-8) were also treated with ethel 48% at the rate of 2 ml L-1 water to break dormancy. A commercial peat-based inoculum of Bradyrhizobium (mixture of strains THA 201 and THA 205; Department of Agriculture, Ministry of Agriculture and Cooperatives, Bangkok, Thailand) was applied to the seed at the time of planting. Three seeds were planted in each pot and the seedlings were then thinned to two plants pot-1 at 14 days after sowing (DAS). Phosphorus fertilizer as triple superphosphate at the rate of 12.12 g P pot-1 and potassium fertilizer as muriate of potash (KCl) at 15.26 g K pot-1 were applied at 14 DAS. Gypsum (CaSO4) at the rate of 153.08 g pot-1 was applied at 40 DAS. Carbofuran (2, 3-dihydro-2, 2-dimethylbenzofuran-7-ylmethylcarbamate 3% granular) was applied at the pod setting stage. Pests and diseases were controlled by weekly applications of carbosulfan (2-3-dihydro-2, 2-dimethylbenzofuran-7-yl (dibutylaminothio) methylcarbamate 20% w/v, water soluble concentrate) at 2.5 L ha-1, methomyl (S-methyl-N-((methylcarbamoyl)oxy) thioacetimidate 40% soluble powder) at 1.0 kg ha-1 and carboxin (5, 6-dihydro-2-methyl-1, 4-oxath-ine-3-carboxanilide 75% wettable powder) at 1.68 kg ha-1.
Prior to planting, uniform water application was supplied to the pots at FC (17.81%) to provide adequate water for germination. In each pot the water was applied in four fractions and the first fraction was given to the soil surface and the remaining fractions were applied through plastic lines were filled into the three holes at 15, 30 and 45 cm below the soil surface. The soil moisture for all pots was maintained at FC until 21 DAS. After 21 DAS, soil water levels were maintained uniformly at field capacity from planting to harvest in well-watered treatment and then soil moisture of stress treatments were allowed to gradually reduce until they reached predetermined levels of 2/3 AW (14.14%) and 1/3 AW (10.47%), respectively. For each treatment, moisture content was maintained uniformly with not lower than 1% moisture change of the predetermined levels until harvest.
In maintaining the specified soil moisture levels, water was added to the respective pots based on crop water requirement and surface evaporation which were calculated following the methods described by Songsri et al. (2009).
Crop water requirement (ETcrop) was calculated as:
| (1) |
where, ETcrop = crop water requirement (mm/day), ETo = evapotranspiration of a reference plant under specified conditions calculated by pan evaporation method, Kc = the crop water requirement coefficient for peanut, which varies with genotype and growth stage.
Surface evaporation (Es) was calculated as:
| (2) |
where, Es = soil evaporation (mm), β = light transmission coefficient measured depending on crop cover, Eo = evaporation from class A pan (mm/day), t = days from the last irrigation or rain (day).
The water supplied to individual pot was equal to the sum of water used by the crop and soil surface evaporation. The calculated amount of water was divided into four fractions. The method of water application was described earlier for uniformity in all columns in the pot.
Data collections
Weather parameters and soil moisture: Weather data for both GH1 and GH2 were obtained from a meteorological station just 50 m away from the greenhouse are presented in Fig. 1a-d. The seasonal mean maximum and minimum air temperature ranged between 33.38 and 21.83°C in GH1 and 34.52 and 24.34°C in GH2. Daily pan evaporation ranged from 2.08 to 9.38 mm in GH1 and 0.10 to 10.14 mm in GH2. The seasonal mean relative humidity 81.27% in GH1 and 90.67% in GH2. The seasonal means of solar radiation were 19.21 MJ m-2 days-1 in GH1 and 15.71 MJ m-2 days-1 in GH2.
Soil moisture were measured by gravimetric method at harvest at the depth of 0-60 cm was presented in Table 1.
Pod yield and harvest index: For each pot, the plants were harvested at maturity and their pods were separated. A sample of shoots was oven-dried at 80°C for 48 h and dry weight was measured. Shoot dry matter content was then calculated to determine shoot dry weight for a pot. Pod yield were weighed after air drying approximately 7-8% moisture content. Harvest index (HI) was calculated as total pod yield/total biomass including pod at final harvest.
Drought tolerance indices (DTI) were calculated for pod yield (DTI (PY)) under stress conditions (2/3 AW or 1/3 AW) to that under well-watered (FC) conditions as suggested by Nautiyal et al. (2002), Songsri et al. (2008a-c) and Songsri et al. (2009), using the relationship as follows:
| (3) |
Number of reproductive parts and yield components: The number of flowers was recorded daily on plants from each pot during the morning (600-800 h, Thailand standard time) from the date of first flowering until harvest. The numbers of reproductive parts were recorded at harvest as the number of pegs (reproductive sink number; RSn = hanging pegs + total pods), number of total pods (immature and mature pods) and number of mature pods per plant (mature pods was separated from immature pods, which were identified by their shriveled seeds and dark internal pericarp color).
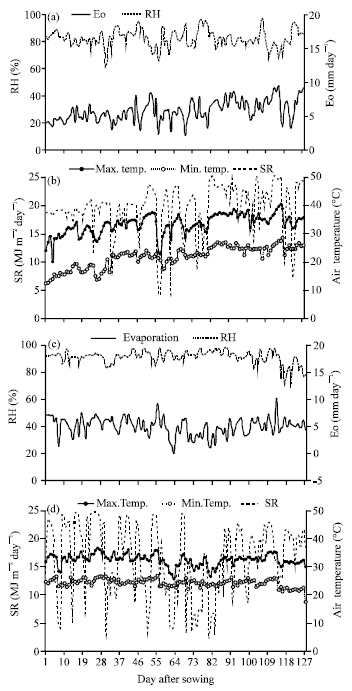 | |
| Fig. 1: | Evaporation (EO), relative humidity (RH), maximum and minimum air temperature (Max and Min Temp.) and solar radiation (SR) in (a, b) GH1 and (c, d) GH2 |
Number of seeds per pod was recorded on the plants in each pot for flower counts at final harvest. Seed size (100 seed weight) was determined from harvested seeds.
Four reproductive efficiency traits (RET) were calculated as follows:
| (4) |
| (5) |
| (6) |
| (7) |
Statistical analysis: Analysis of variance was performed individually for each character in each experiment. Error variances for the two seasons were tested for homogeneity by Bartlett’s test (Hoshmand, 2006). Combined analyses of variance were done for those characters that error variances for the two seasons were homogeneous and Duncan’s Multiple Range Test (DMRT) was used to compare means.
Multiple-linear regression was used to determine the relative contribution of yield component or reproductive efficiency to pod yield under FC, 2/3 AW and 1/3 AW. The analysis was based on the following statistical model (Hoshmand, 2006):
| (8) |
where, Yi is pod yield of genotype i, α is the Y intercept, X1i, X2i and Xni are yield components or reproductive efficiency of genotype i, respectively, β1, β2 and β3 are regression coefficients for the independent variables X1, X2 and X3 and δi is the associated deviation from regression.
The analysis was carried out by fitting the full model first and then determining the relative importance of the individual independent variables. A sequential fit was then performed by fitting the more important variable first.
The relative contributions of the individual independent variables to pod yield under FC, 2/3 AW and 1/3 AW were determined from the percentages of regression sum of squares due to the respective independent variables to total sum of squares in the sequential fitted analysis.
RESULTS AND DISCUSSION
Soil moisture level was monitored using gravimetric method (Table 1). The results showed reasonably good management of soil moistures. Observations found visual wilting in 2/3 AW and more severe wilting in 1/3 AW in the afternoon. RWC of the plants in treatment 1/3 AW were extremely low at 97 DAS (data not presented).
Combined analysis of variance: Combined analysis of variance showed a significant (p≤0.01) differences among the water regimes and genotypes for all traits (Table 2). The difference in seasons was also significant for seed per pod, number of flowers, reproductive sink per plant, RET 2 and RET 4 (p≤0.01) and 100 seed weight (p≤0.05). The WxG interaction effects were significant for RET 2 (p≤0.05) and each other (p≤0.01).
| Table 1: | Soil moisture content at 0-60 cm at the end of stress period during each water regimes |
 | |
| Soil moisture of FC = 17.81%, 2/3AW = 14.14%, 1/3AW = 10.47% and PWP = 6.80%; Soil moisture content of theoretical field capacity (-0.01 MPa) and permanent wilting point (-1.5 MPa) was determined by the pressure plate method | |
| Table 2: | Mean squares from the combined ANOVA for pod yield (PY), harvest index (HI), 100 seed weight, number of seed per pod, number of mature pod, flowers, reproductive sink and pods per plant and percentage of flowers to reproductive sink (RET 1), reproductive sink to pods (RET 2), pods to mature pods (RET 3) and flowers to mature pods (RET 4) at harvest under three water regimes of 11 genotypes in GH1 and GH2, 2002/03 |
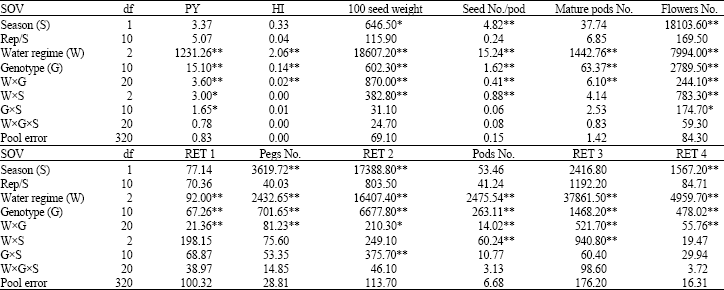 | |
| * and **Significant at p = 0.05 and significant at p = 0.01, respectively. df: Degree of freedom, SOV: Source of variation | |
| Table 3: | Pod yield (g plant-1) and drought tolerance index (DTI) for 11 peanut genotypes grown under different water regimes at harvest |
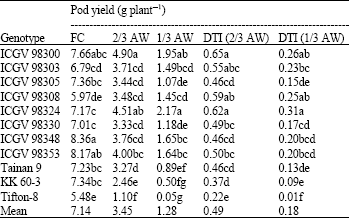 | |
| Mean in the same column with the same letters are not significantly different by Duncan’s multiple range test (DMRT) (at p≤0.05). DTI for a genotype were calculated by the ratio of stressed (2/3 available water (AW) or 1/3 AW)/ non-stressed (field capacity; FC) conditions | |
The SxW interaction effects were significant for PY (p≤0.05) and each other (p≤0.01). For GxS interactions were significant for RET 2 and RET 3 (p≤0.01) and pod yield and number of flower per plant (p≤0.05). WxGxS interactions were not significant for all traits.
Pod yield and drought tolerance index: Drought significantly reduced pod yield by 51% at 2/3 AW and 82% at 1/3 AW conditions (Table 3). ICGV 98348 and ICGV 98353 had high pod yield under FC. ICGV 98300 had high pod yield under both well-watered and drought conditions. The yield stabilizing strategy for this genotype should be largely due to their high starting yield at FC and in minor part due to their relatively low reductions (high DTI). ICGV 98324 exhibited high pod yield under drought conditions because of its high DTI (low yield reduction). Low reduction in pod yield was more important for stabilizing the yield of these genotypes under drought. The genotypes with poor performance for pod yield under both non-stressed and stressed conditions were Tifton-8 and KK 60-3. Tiffton-8 had the lowest stating point for yield (yield potential). The reductions in pod yield of these genotypes were also relatively high, indicating that they were most sensitive to drought.
Two possible strategies of drought resistance may be useful for explaining drought resistance in these peanut genotypes. Genotypes with high pod yield under drought conditions should be of either (1) high pod yield under well-watered conditions (e.g., ICGV 98300) or (2) ability to maintain a low rate of yield reduction under increasing stressed (high DTI) (e.g., ICGV 98324 and ICGV 98300).
| Table 4: | Harvest index (HI), number of mature pods per plant, 100 seed weight (g) and number of seed per pod of 11 peanut genotypes grown under different water regimes at harvest |
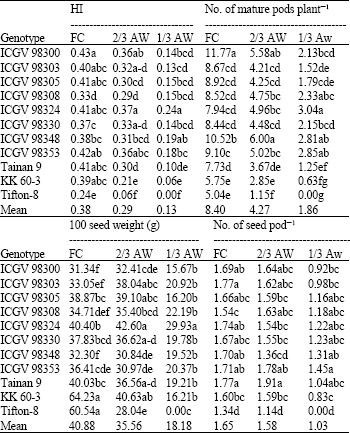 | |
| Mean in the same column with the same letters are not significantly different by Duncan’s multiple range test (DMRT) (at p≤0.05) | |
HI and yield components: Water stress reduced the HI, number of mature pods, seeds per pod and seed size (Table 4). The genotype with high HI under both non-stressed and stressed conditions (2/3 AW and 1/3 AW) was ICGV 98324. ICGV 98300, ICGV 98303 and ICGV 98353 had high HI under FC and 2/3 AW conditions. Tifton-8 had the lowest HI at all water regimes.
ICGV 98300, ICGV 98348 and ICGV 98324 had the highest number of mature pods per plant under FC, 2/3 AW and 1/3 AW, respectively.
Tifton-8 and KK 60-3 had the largest seed size under well-watered, but sharply decreased when available soil water levels were reduced (Table 4). ICGV 98324 had the largest seed size under both mild and severe droughts.
There were quite small differences among peanut genotypes for number of seeds per pod at any water levels. Tainan 9 and ICGV 98353 showed high numbers of seeds per pod and Tifton-8 had the lowest number of seeds per pod under all conditions (Table 4). Although, the genetic variation was low, clearer differences were observed under drought conditions, indicating differences in drought sensitivity among these peanut genotypes for number of seeds per pods.
| Table 5: | Contributions of yield components to pod yield under FC, 2/3AW and 1/3 AW conditions |
 | |
| * and **Significant at p≤0.05 and p≤0.01, respectively | |
KK 60-3 and Tifton-8 had high seed size under non-stressed conditions. These Virginia type large-seeded peanuts also had higher rates of reduction in seed size than did the medium-seeded genotypes under water stressed conditions. The rate of reduction in seed size of Tifton-8 was higher than that of KK 60-3, making it perform poorer under water stressed conditions. ICGV 98324 was not among the largest under non-stressed conditions, but under stressed conditions it had seed size larger than others.
Multiple regression analysis showed the contributions to pod yield of number of mature pods per plant, seed size and number of seed per pod under FC, 2/3 AW and 1/3 AW conditions (Table 5). The contribution of number of mature pods per plant was highest under mild and severe drought stress (82.70 and 89.22%, respectively), whereas the contribution of number of mature pods per plant and seed per pod were less under FC conditions (27.94 and 26.66%, respectively). Number of mature pods per plant seemed to play an important role in maintaining high pod yield under both mild and severe stressed conditions. The contributions of seed size to pod yield were highest under well-watered (41.83%) but the contribution seemed to be reduced under mild and severe drought stress conditions (8.88 and 5.96%, respectively).
Reproductive traits and reproductive efficiency
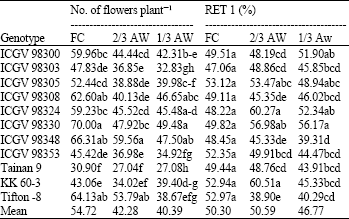
Flowers number and its conversion to reproductive sink: Number of flowers per plant and percentage of flowers to reproductive sink (percent fertilization) was reduced under severe drought condition (Table 6). ICGV 98330 had the highest number of flowers per plant under FC and 1/3 AW and ICGV 98348 under 2/3 AW. Tainan 9 had the lowest flower number per plant at all water regimes.
Variability of peanut genotypes for percentage of flowers to reproductive sink (RET 1) was found significant under drought stress conditions. ICGV 98324, ICGV 98330 and ICGV 98305 had high RET 1 under 2/3 AW and 1/3 AW. KK 60-3 and ICGV 98300 were also high percent fertilization under 2/3 AW and 1/3 AW, respectively. ICGV 98348 and Tifton-8 had low percentage of flowers to reproductive sink under drought conditions (Table 6).
| Table 6: | Number of reproductive sink and percentage of flower to reproductive sink (RET 1) of 11 peanut genotypes grown under different water regimes at harvest |
 | |
| Mean in the same column with the same letters are not significantly different by Duncan’s multiple range test (DMRT) (at p≤0.05) | |
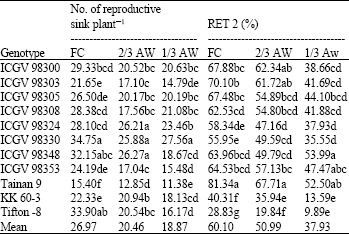
| Table 7: | Number of reproductive sink per plant and percentage reproductive sink to pods (RET 2) of 11 peanut genotypes grown under different water regimes at harvest |
 | |
| Mean in the same column with the same letters are not significantly different by Duncan’s multiple range test (DMRT) (at p≤0.05) | |
Reproductive sink number and its conversion to pods: Reproductive sink number and percentage of reproductive sink to pods (RET 2) were decreased with droughts (Table 7). ICGV 98330 had the highest number of reproductive sink per plant at all water regimes (34.75, 25.88 and 27.56 under FC, 2/3 AW and 1/3 AW, respectively). Tainan 9 had the lowest reproductive sink per plant at all water regimes. In contrast, Tainan 9 had high RET 2 at all FC, 2/3 AW and (81.34, 67.71 and 52.50% under FC, 2/3 AW and 1/3 AW, respectively). ICGV 98348 had the highest RET 2 at 1/3 AW (53.99%). Tifton-8 had the lowest RET 2 at all water regimes (28.83, 19.84 and 9.89% under FC, 2/3 AW and 1/3 AW, respectively).
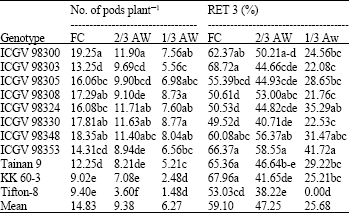
Pods number and its conversion to mature pods: Number of pods per plant and percentage of pods to mature pods (RET 3) were decreased with droughts (Table 8). ICGV 98300 had high number of pods per plant at all water regimes (19.25, 11.90 and 7.56 under FC, 2/3 AW and 1/3 AW, respectively).
| Table 8: | Number of pods per plant and percentage pods to mature pods (RET 3) of 11 peanut genotypes grown under different water regimes at harvest |
 | |
| Mean in the same column with the same letters are not significantly different by Duncan’s multiple range test (DMRT) (at p≤0.05) | |
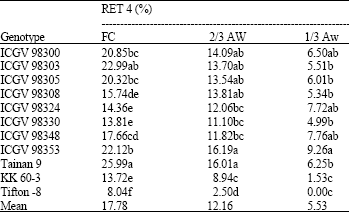
| Table 9: | Percentage of flowers to mature pods (RET 4) of 11 peanut genotypes grown under different water regimes at harvest |
 | |
| Mean in the same column with the same letters are not significantly different by Duncan’s multiple range test (DMRT) (at p≤0.05) | |
ICGV 98308 and ICGV 98330 also had high number of pods per plant under severe drought condition (8.73 and 8.77, respectively). Tifton-8 and KK 60-3 had low reproductive sink per plant at all water regimes. ICGV 98353 had high RET 3 under FC, 2/3 AW and 1/3 AW (66.37, 58.55 and 41.72%, respectively). ICGV 98330 had the lowest RET 3 under well-watered condition (49.52%). Tifton-8 had the lowest under both mild and severe drought conditions (38.22 and 0.00%, respectively).
Conversion of flowers to mature pods: Percentage of flowers to mature pods (RET 4) was decreased with droughts (Table 9). The conversion of flowers to mature pods of peanut genotypes under FC, 2/3 AW and 1/3 AW were 17.78, 12.16 and 5.53%, respectively. The ability of peanut genotypes to conversing flowers to mature pods was different.
Tainan 9 had the highest RET 4 under well-watered condition (25.99%). ICGV 98353 had the highest RET 4 at 2/3 AW and 1/3 AW (16.19 and 9.26%, respectively). ICGV 98300 also high RET 4 under mild and severe drought conditions. Tifton-8 had the lowest under FC, 2/3 AW and 1/3 AW conditions (8.04, 2.50 and 0.00%, respectively).
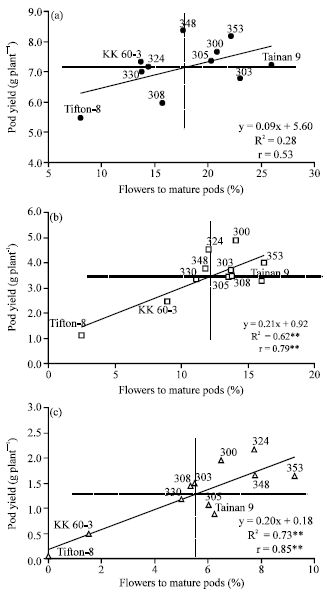 | |
| Fig. 2: | Relationship between flower to mature pods (%) and pod yield (g plant-1) under FC (a), 2/3 AW (b) and 1/3 AW (c) of 11 peanut genotypes. **Significant at p≤0.01 |
The ability to transform flowers to mature pods had strong and positive correlation to pod yield under 2/3 AW and 1/3 AW (0.79 and 0.85, respectively) (Fig. 2a-c). The peanut genotypes with high flower production at FC (Tifton-8) did not produce high number of pegs and pods under stress, but some genotypes with lower flower production at FC (ICGV 98324) could produce similar numbers of pegs and pods under drought. This might indicate that flower production of peanut is generally sufficient for attaining acceptable yield even under drought conditions.
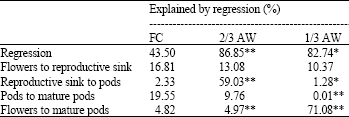
| Table 10: | Contributions of reproductive traits to pod yield under FC, 2/3AW and 1/3 AW conditions |
 | |
| * and **Significant at p≤0.05 and p≤0.01, respectively | |
Pegging and pod set responses of various peanut genotypes under drought varied substantially, leading to large differences in pod yield and the reductions in pod yield also varied among peanut genotypes (Nageswara Rao et al., 1989; Songsri et al., 2008c). However, the genotypes producing the lowest number of flowers under normal conditions rarely produced high pod yield under water stress (Songsri et al., 2008c). The peanut genotypes with extremely low flower production are therefore inferior to those with abundant flowers under drought conditions. However, in a parallel study, Songsri et al. (2008c) did not provide the information in term of reproductive efficiency that contributing to pod yield. The results in this study indicated that the line ICGV 98324 with high reproductive efficiency had high HI and pod yield under severe drought conditions, and, under moderate drought conditions (2/3 AW), ICGV 98300 had high RET 4. In previous study, ICGV 98324 and ICGV 98300 were identified as drought resistant lines based on low specific leaf area (leaf thickness) and large root system, respectively (Songsri et al., 2009). The drought resistant trait in each genotype might contribute to reproductive efficiency and ultimately contribute to pod yield.
Contribution of Reproductive Efficiency Traits (RET) to pod yield: Multiple regression analysis showed the contributions of RET to pod yield under FC, 2/3 AW and 1/3 AW conditions (Table 10). The contribution of the conversion of reproductive sink to pods (RET 2) was highest under mild drought stress (59.03%).
The contribution of the conversion of flower to mature pod (RET 4) was highest under severe drought stress conditions (71.08%).
The result indicated that under mild drought stress (2/3 AW) the reproductive phase between after flower pollination to poding stage may be the most critical period to maintain high pod yield. The critical period was wider under more severe drought (1/3 AW). Under severe conditions, all reproductive stages from flowering to fulfilled mature pod were critical in maintaining high yield.
CONCLUSION
High number of mature pods is advantageous and necessary for high yield under drought conditions. The maintaining high pod yield under drought should be due to high potential yield and/or low reduction yield. In drought resistant genotypes, the ability of reproductive parts to converse to mature pod plays an important role in maintaining yield under drought. This information provides a better understanding on the factors contributing to higher pod yield under drought and assists peanut breeders to formulate more effective and efficient breeding strategies for improving drought resistance in peanut.
ACKNOWLEDGMENTS
The authors are grateful for the financial support of the Royal Golden Jubilee Ph.D. Program (Grant No. PHD/0190/2544), the Senior Research Scholar Project of Professor Dr. Aran Patanothai under the Thailand Research Fund and the Senior Research Scholar Project of Assoc. Prof. Dr. Sanun Jogloy under Khon Kaen University. We thank the work of many people for data collection and processing.
REFERENCES
- Awal, M.A. and T. Ikeda, 2002. Recovery strategy following the imposition of episodic soil moisture deficit stands of peanut (Arachis hypogaea L.). J. Agron. Crop Sci., 188: 185-192.
Direct Link - Chapman, S.C., M.M. Ludlow, F.P.C. Blamey and K.S. Fischer, 1993. Effect of drought during early reproductive development on the dynamics of yield development of genotypes of peanut (Arachis hypogaea L.) II. Biomass production, pod development and yield. Field Crops Res., 32: 211-225.
CrossRefDirect Link - Coffelt, T.A., R.O. Hammons, W.D. Branch, R.W. Mozingo and P.M. Phipps et al., 1985. Registration of Tifton-8 peanut germplasm. Crop Sci., 25: 203-203.
Direct Link - Holbrook, C.C. and H.T. Stalker, 2003. Peanut breeding and genetic resources. Plant Breed. Rev., 22: 297-356.
Direct Link - Hoshmand, A.R., 2006. Design of Experiments for Agriculture and the Natural Sciences. 2nd Edn., Chapman and Hall, New York.
Direct Link - Nageswara Rao, R.C., S. Singh, M.V.K. Sivakumar, K.L. Srivastava and J.H. William, 1988. Effect of water deficit at different growth phase of peanut. II yield response. Agron. J., 80: 431-438.
Direct Link - Nageswara Rao, R.C., J.H. Williams and M. Singh, 1989. Genotypic sensitivity to drought and yield potential of peanut. Agron. J., 81: 887-893.
Direct Link - Nautiyal, P.C., V. Ravinda, P.V. Zala and Y.C. Joshi, 1999. Enhancement of yield in peanut following the imposition of transient soil-moisture-deficit stress during the vegetative phase. Exp. Agric., 35: 371-385.
CrossRefDirect Link - Nautiyal, P.C., R.C. Nageswara Rao and Y.C. Joshi, 2002. Moisture-deficit-induced changes in leaf-water content, leaf carbon exchange rate and biomass production in groundnut cultivars differing in specific leaf area. Field Crops Res., 74: 67-79.
CrossRefDirect Link - Nigam, S.N., M.S. Basu and A.W. Cruickshank, 2003. Hybridization and description of the trait-based and empirical selection programs. Proceedings of the International Workshop on Breeding for Drought-Resistant Peanuts, February 25-27, 2003, ICRISAT Centre Andhra Pradesh, India, ACIAR, Canberra, Australia, pp: 15-17.
Direct Link - Nigam, S.N., S. Chandra, K.R. Sridevi, M. Bhukta and A.G.S. Reddy et al., 2005. Efficiency of physiological trait-based and empirical selection approaches for drought tolerance in groundnut. Ann. Applied Biol., 146: 433-439.
CrossRefDirect Link - Rachaputi, N.R., G.C. Wright and S. Krosch, 2002. Management practices to minimise pre-harvest aflatoxin contamination in Australian peanuts. Aust. J. Exp. Agric., 42: 595-605.
CrossRefDirect Link - Reddy, T.Y., V.R. Reddy and V. Anbumozhi, 2003. Physiological responses of peanut (Arachis hypogaea L.) to drought stress and its amelioration: A critical review. Plant Growth Regul., 41: 75-88.
Direct Link - Songsri, P., S. Jogloy, N. Vorasoot, C. Akkasaeng, A. Patanothai and C.C. Holbrook, 2008. Root distribution of drought-resistant peanut genotypes in response to drought. J. Agron. Crop Sci., 194: 92-103.
CrossRefDirect Link - Songsri, P., S. Jogloy, T. Kesmala, N. Vorasoot, C. Akkasaeng, A. Patanothai and C.C. Holbrook, 2008. Heritability of drought-resistance traits and genotypic an phenotypic correlation of drought-resistance and agronomic traits in peanut. Crop Sci., 48: 2245-2253.
Direct Link - Songsri, P., S. Jogloy, T. Kesmala, N. Vorasoot, C. Akkasaeng, A. Patanothai and C.C. Holbrook, 2008. Responses of reproductive characters of drought resistant peanut genotypes to drought. Asian J. Plant Sci., 7: 427-439.
Direct Link - Songsri, P., S. Jogloy, C.C. Holbrook, T. Kesmala, N. Vorasoot, C. Akkasaeng and A. Patanothai, 2009. Association of root, specific leaf area and SPAD chlorophyll meter reading to water use efficiency of peanut under different available soil water. Agric. Water Manage., 96: 790-798.
CrossRefDirect Link - Vorasoot, N., P. Songsri, C. Akkasaeng, S. Jogloy and A. Patanothai, 2003. Effect of water stress on yield and agronomic characters of peanut (Arachis hypogaea L.). Songklanakarin J. Sci. Technol., 25: 283-288.
Direct Link - Vorasoot, N., C. Akkasaeng, P. Songsri, S. Jogloy and A. Patanothai, 2004. Effect of available soil water on leaf development and dry matter partitioning in 4 cultivars of peanut (Arachis hypogaea L.). Songklanakarin J. Sci. Technol., 26: 787-794.
Direct Link