
M. Niakan
Department of Biology, Islamic Azad University, Gorgan Branch, Iran
K. Saberi
Department of Biology, Islamic Azad University, Gorgan Branch, Iran
Asian Journal of Plant Sciences
Year: 2009 | Volume: 8 | Issue: 6 | Page No.: 440-446







ABSTRACT
In this study, different amounts of aqueous extracts [0, 5, 15 and 30% (v/v)] and decompose of Eucalyptus leaves [(0, 3 and 6% (w/w)] were applied to Phalaris in pot culture and evaluated on growth parameters and antioxidant enzymes activity. The results showed that most of the growth parameters of phalaris were decreased when exposed to different amounts of decompose and water extracts of Eucalyptus leaves, especially in the leaf decompose. Also, antioxidant enzymes activity in root and shoot of Phalaris were affected by aqueous extracts and decompose of Eucalyptus leaves. The findings indicated that decompose of Eucalyptus leaves decreased catalase activity and increased ascorbate activity in root and shoot of Phalaris in comparison to control while aqueous extract of Eucalyptus leaves significant raised ascorbate peroxidase in Phalaris root. Activity of other enzymes such as polyphenol oxidase and peroxidase in Phalaris affected less aqueous extracts and decompose of Eucalyptus leaves.
PDF Abstract XML References Citation
How to cite this article
M. Niakan and K. Saberi, 2009. Effects of Eucalyptus Allelopathy on Growth Characters and Antioxidant Enzymes Activity in Phalaris Weed. Asian Journal of Plant Sciences, 8: 440-446.
DOI: 10.3923/ajps.2009.440.446
URL: https://scialert.net/abstract/?doi=ajps.2009.440.446
DOI: 10.3923/ajps.2009.440.446
URL: https://scialert.net/abstract/?doi=ajps.2009.440.446
INTRODUCTION
In classic opinion, allelopathy refers to the chemical inhibition of one species by another. Modern research suggested that allelopathic effects can be both positive and negative, depending upon the doses and organisms (Bais et al., 2003). When plants are exposed to allelochemicals, their growth and development are affected (Putnam and Duke, 1978; Niakan et al., 2008). The readily visible effects include inhibited or retarded germination rate, seeds darkened and swollen, reduced root or radicle and shoot or coleoptile extension, swelling or necrosis of root tips, curling of the root axis, discoloration, lack of root hairs, increased number of seminal roots, reduced dry weight and lowered reproductive capacity. These gross morphological effects may be secondary manifestations of primary events, caused by a variety of more specific effects acting at the cellular or molecular level in the receiver plants (Singh et al., 2006; Peng et al., 2004; Prati and Bossdorf, 2004). The activity of enzymes was also affected by allelopathic compounds (Peng et al., 2004; Niakan et al., 2008). Environmental stress can create oxidative stress through increased production of Reactive Oxygen Species (ROS), e.g., singlet oxygen (1O2), hydroxyl radical (HO°) and hydrogen peroxide (H2O2) (Blokhina et al., 2003; Lee et al., 2007; Gechev and Hille, 2005; Apel and Hirt, 2004). Reactive oxygen species may cause lipid peroxidation whereas superoxide dismutase (SOD) and peroxidase (POD) are inhibiting (Blokhina et al., 2003; Singh et al., 2006). Under oxidative stress, plants respond by increasing antioxidant defenses, notably enzymes such as SOD and POD. However, excessive ROS may cause a decrease in SOD and POD activities (Mishra et al., 1993). Numerous studies have supported the signaling role of reactive oxygen species during different environmental., responses and developmental processes, including biotic and abiotic stress responses, allelopathic plant-plant interactions, cell division and elongation and programmed cell death (Apel and Hirt, 2004; Bais et al., 2003; Pei et al., 2000).
Eucalyptus is one of the most important tree species for wood production in the world. It is said that Eucalyptus is toxic, due to allelopathic properties, which serve to reduce germination of other plant (Khan et al., 2005; Shiming, 2005).
Batish et al. (2004) reported that Eucalyptus citriodora produces volatile materials that inhibit the germination, seedling length, chlorophyll content and respiratory ability in Phalaris minor, Chenopodium album, Echinochola crus-gali, Ageratum conyzoide, Parthenium hysterphorus and Amaranthus sp. Shiming (2005) showed the allelopathic effects of 4 species of Eucalyptus on radish seed germination and seedling growth.
Researches showed that Eucalyptus species released volatile compounds such as benzoic, cinnamic and phenolic acids, which inhibited growth of crops and weeds growing near it (Sasikumar et al., 2001; Kohli et al., 1998). Daizy et al. (2004) reported that volatile oil from leaves of Eucalyptus citriodora significantly reduced germination seedling length and chlorophyll content in Triticum aestivum, Zea mays and Raphanus sativus. Batish et al. (2004) reported that water extracts of E. citriodora significantly reduced weed establishment.
As a scientific discipline, allelopathy is still relatively new. But it has already contributed to the solution of practical problems in agriculture and provided explanations for observed plant-plant interactions. Allelochemicals might be directly used as natural product pesticides as well as models for developing new pesticides. By modifying these allelochemicals, the end product could be more active, selective, or persistent.
The aim of the present study was to determine the inhibitory effect of decompose and aqueous extract of Eucalyptus leaf on growth, generation of reactive oxygen species, as indicators of oxidative stress and changes in activities of antioxidant enzymes in Phalaris weed.
MATERIALS AND METHODS
In summer 2008, Eucalyptus leaves (Eucalyptus camaldulensis) were harvested from natural habitat located on Azad shahr city (37.053 longitude, 55.095 latitude and 34 m above sea level) in North Iran. Then leaves were dried in shade and grinded. Dried samples were mixed with soil (Si-Clay tissue) in ratio (0, control), 3 and 6% (w/w) and this mixture were placed in shade for 30 days.
Aqueous extract of Eucalyptus leaves preparation: Five grams of dried Eucalyptus leaves (Eucalyptus camaldulensis) were added to 150 mL distilled water and were shaked for 12 h. The mixture was passed through Wathman paper (number 2) and filtrated through microporous filter (0.2 μm pore side) (Narwal and Tauro, 1996) and prepared aqueous extract in concentrations 0, 5, 15 and 30% (v/v).
Phalaris planting
Phalaris planting in Decompose of Eucalyptus leaves: Phalaris seeds (Phalaris minor) were imbibed in distilled water for 48 h and then divided to 2 parts. The first part was planted in pots that include mixture of soil (Si-Clay tissue) and Eucalyptus Eucalypt leaves in ratio 0 (control), 3 and 6%. Pots maintained in a photoperiod of 10-14 (L-D) and controlled temperature (20±2°C) and irrigated 3 days interval with 200 mL water. After 60 days plants were harvested and used for growth parameters measurement and biochemical assays.
Phalaris planting for spray aqueous extract of Eucalyptus leaves: The Phalaris seeds were placed in water for 48 h and planted in pots soils with Si-Clay tissue. After 30 days, Phalaris plants were sprayed to aqueous extract of Eucalyptus leave in concentrations of 0 (control), 5, 15 and 30% (v/v) for 3 times in week during 30 days. Then, plants were brought out of soil and used for growth parameters measurement and biochemical assays.
Growth parameters measurement: After 60 days, root and shoot length of Phalaris plants that had been treated by decompose and aqueous extract of Eucalyptus leaves were measured. Root and shoot length, fresh and dry weights of root and shoot and leaf area of Phalaris plants under treatments of decompose and aqueous extract of Eucalyptus leaves also were determined. For dry weight, root and shoot of Phalaris were placed in hot-air oven at 90°C for 24 h.
Biochemical assays
Enzyme extraction: one gram of the roots or shoots of Phalaris plants treated with decompose or aqueous extract of Eucalyptus leaf were weighted and homogenized with 4 mL extract solution containing 1.2 g Tries, 2 g ascorbate, 3.8 g borax (Di-sodium tetra borate), 2g EDTA-Na2, 50 g polyethylene glycol 2000 in 100 mL distilled water. The solution was placed in 4°C for 24 h and then was centrifuged for 30 min at 4000 g. The clear supernatant was taken as enzyme extract and used of them for catalase and peroxidase, poly phenol oxidase and ascorbate peroxidase activity assay.
Catalase activity assay: The catalase activity was assayed by Chance and Maehly (1955) method with the following modification: 5 mL of assay mixture for catalase activity contained: 300 μM of phosphate buffer (pH 6.8), 100 μM of H2O2 and 1 mL of the twice diluted enzyme extracted. After incubation at 25°C for 1 min, the reaction was stopped by adding 10 mL of 2% (v/v) H2SO4 and the residual H2O2 was titrated against 0.01 N of KMnO4 until a faint purple color persisted for at least 15 sec. A control was run at the same time but the enzyme activity was stopped at zero time. One unit of catalase activity is defined as the amount of enzyme which breaks down 1 μmol of H2O2 min-1 under the described assay condition.
Peroxidase activity assay: The peroxidase activity was determined by Koroi (1989) method. The 0.1 mL of enzyme extract was added to assay mixture containing: 2 mL 0.2 M acetate buffers (pH 5.0), 0.4 mL of 3% H2O2 and 0.2 mL of 0.01 M benzidin solution in 50% alcohol. The absorbance was taken at 530 nm. In order to protect of enzyme activity, the experiment was done in chilled conditions.
Ascorbate peroxidase activity assay: Assay mixture of ascorbate peroxidase activity was consisted of: 2 mL phosphate buffer 0.05 M (pH 6.5) were mixed to 0.2 mL H2O2 3%, 0.2 mL ascorbic acid 50 μM and 0.1 mL enzyme extract was added to them. The enzyme activity was determined by taking the absorbance at 265 nm according (Arrigoni, 1994).
Poly phenol oxidase activity assay: The activity of poly phenol oxidase was assayed by Manoranjan and Mishra (1976) Assay mixture was consisted of: 1.5 mL of 0.2 M phosphate buffer (pH 7.6), 0.4 mL pyrogallol 0.02 M and 0.1 mL enzyme extract. This mixture was incubated at 28°C for 3 min. The enzyme activity was determined by measuring the absorbance at 430 nm spectrophotometrically.
Statistical analysis: The experiment was carried out in completely randomized design with 4 replications The statistical significance of the difference between parameters was evaluated by means of Duncan-test on SPSS 11.5 and for each treatment and control 4 replications was selected. The results are given in the text as p, the probability values and p≤0.01 was adopted as criterion of significance.
RESULTS
The results of this study showed that by increasing amount of decompose Eucalyptus leaf in soil (3, 6%) length, fresh and dry weight of Phalaris root and shoot at p≤0.01 decrease (Table 1). Also, application of high concentrations of aqueous extract of Eucalyptus leaf (15, 30%) decreased fresh and dry weight of Phalaris root in comparison to control and 5% aqueous extract but they had a non significant effect on fresh and dry weight of Phalaris shoot at p≤0.01 (Table 2).
Present results also indicated that root length of Phalaris plants treated with 5, 15 and 30% concentrations of aqueous extract of Eucalyptus leaf were more than control while shoot length of Phalaris by increasing concentration of Eucalyptus leaf extract raised significantly (p≤0.01, Table 2).
Statistical analysis of the data showed that treatment with Eucalyptus leaf decompose decreased Phalaris leaf area more than spray of Eucalyptus leaf aqueous extract (Table 3).
| Table 1: | Effects of different amounts of decompose of Eucalyptus leaf (in each ratio 0: Control, 3, 6% with soil) on root and shoot length (cm), dry and fresh shoot and root (g) of Phalaris |
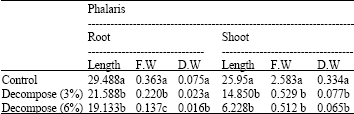 | |
| Means±SD followed by different letter(s) of the same column indicates that the values are significant different at 0.01 determined by ANOVA and Duncan multiple range tests | |
| Table 2: | Effects of different amounts of aqueous extracts (0, 5,15 and 30%) of Eucalyptus leaf on root and shoot length (cm), dry and fresh shoot and root (g) of Phalaris |
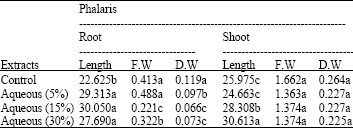 | |
Means±SD followed by different letter(s) of the same column indicates that the values are significant different at 0.01 determined by ANOVA and Duncan multiple range tests | |
| Table 3: | Effects of different amounts of decompose of Eucalyptus leaf in ratio 0: Control, 3 and 6% with soil and aqueous extracts (0, 5, 15 and 30%) of Eucalyptus leaf on leaf area (cm2) of Phalaris |
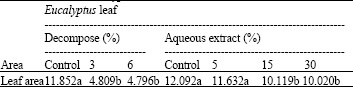 | |
| Means±SD followed by different letter(s) of the same column indicates that the values are significant different at 0.01 determined by ANOVA and Duncan multiple range tests | |
Also, the analysis of data revealed that decompose of Eucalyptus leaf had more inhibition effect on fresh and dry weight of root and shoot of Phalaris than aqueous extract of Eucalyptus leaf (Table 1, 2).
These findings reveal that most of the growth parameters of Phalaris were decreased when exposed to different amounts of decompose and water extracts of Eucalyptus leaf. The reduction is more severe by Eucalyptus leaf decompose.
Table 4-7 show antioxidant enzymes activity in leaf and root of Phalaris.
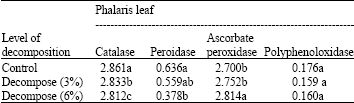
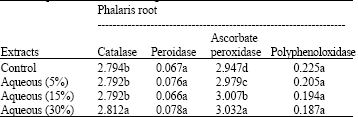
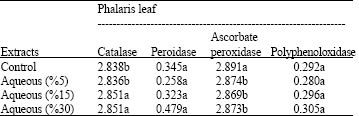
Catalase activity in leaves and roots of Phalaris decreased by increasing the amount of Eucalyptus leaf decompose (Table 4, 5) while its activity in leaves and roots was increased at higher concentration of aqueous extract of Eucalyptus leaf (Table 6, 7).
At highest content of Eucalyptus leaf decompose (6%) peroxidase activity in Phalaris leaf decreased (Table 6). Also, significant difference in peroxidase activity of Phalaris roots between control and several amounts of Eucalyptus leaf decompose (3, 6%) was not observed (Table 4).
| Table 4: | Effects of different amounts of decompose of Eucalyptus leaf (in ratio 0: Control 3, 6% with soil) on antioxidant enzyme activity on Phalaris root include: catalase, peroxidase, poly phenol oxidase and ascorbate peroxidase (OD min-1 g-1 Fw) |
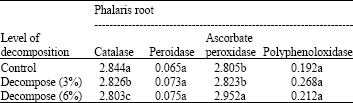 | |
| Means±SD followed by different letter(s) of the same column indicates that the values are significant different at 0.01 determined by ANOVA and Duncan multiple range tests | |
| Table 5: | Effects of different amounts of decompose of Eucalyptus leaf (in ratio 0: Control, 3, 6% with soil) on antioxidant enzyme activity on Phalaris leaf include: catalase, peroxidase, poly phenol oxidase and ascorbate peroxidase (OD min-1 g-1 Fw) |
 | |
| Means±SD followed by different letter(s) of the same column indicates that the values are significant different at 0.01 determined by ANOVA and Duncan multiple range tests | |
| Table 6: | Effects of different amounts of aqueous extracts (0, 5, 15 and 30%) of Eucalyptus leaf on antioxidant enzyme activity on Phalaris root include: catalase, peroxidase, poly phenol oxidase and ascorbate peroxidase (OD min-1 g-1 Fw) |
 | |
| Means±SD followed by different letter(s) of the same column indicates that the values are significant different at 0.01 determined by ANOVA and Duncan multiple range tests | |
| Table 7: | Effects of different amounts of aqueous extracts (0, 5,15 and 30%) of Eucalyptus leaf on antioxidant enzyme activity on Phalaris leaf include: catalase, peroxidase, poly phenol oxidase and ascorbate peroxidase (OD min-1 g-1 Fw) |
 | |
| Means±SD followed by different letter(s) of the same column indicates that the values are significant different at 0.01 determined by ANOVA and Duncan multiple range tests | |
Decompose and aqueous extract of Eucalyptus leaf was not significant effect on poly phenol oxidase activity in leaves and roots of Phalaris in compared to control (Table 4-7).
The data indicated that by increasing amount of Eucalyptus leaf decompose ascorbate peroxidase activity enhanced in Phalaris roots and leaves. The increase was significant at 6% concentration (p≤0.01, Table 4, 5).
The results also showed that increasing concentration of Eucalyptus leaf extracts significantly increased ascorbate peroxidase activity in Phalaris roots (Table 6). In Phalaris leaves, the higest ascorbate peroxidase activity was observed in control while a non significant difference was observed in the different concentrations of Eucalyptus leaf extracts (Table 7).
DISCUSSION
The results presented in Table 1 showed that increasing the amount of decompose Eucalyptus leaf in soil decreased length, fresh and dry weight of Phalaris of Phalaris root and shoot. Also, application of highest concentrations of aqueous extract of Eucalyptus leaf (15, 30%) decreased fresh and dry weight in Phalaris root in comparison to control and 5% concentration Shahid et al. (2006) reported that aqueous extract of Eucalyptus camaldulensis showed allelopathic effect on the growth of wheat and its weeds including weeds like Avena fatua, Convolvulus arvensis, Rumex dentatus and Phalaris minor.
Under laboratory conditions, the Enterolobium contortisiliquum plants showed tolerance to Eucalyptus. grandis oil, whereas E. camaldulensis oil caused loss of leaves, inhibition of height and diameter growth and a concomitant decrease in effective PSII quantum yield and the reduction of photosynthetic electron-transport chains (Duarte et al., 2006).
The roots of plants exposed to allelochemicals became brownish, stunted and void of root hairs. This might be due to rapid inhibiting effect on respiration of root tips, which ultimately reduced elongation (Shahid et al., 2006).
Present results also indicated that root and shoot lengths of Phalaris resulting from application of 5, 15 and 30% concentrations of aqueous extract of Eucalyptus leaf were significantly increased. Singh et al. (2006) reported that biological activities of receiver plants to allelochemicals are known to be concentration dependent with a response threshold. Responses are, characteristically, stimulation at low concentrations of allelochemicals and inhibition as the concentration increases. Similar results were reported by Peng et al. (2004). It seems that concentration of Eucalyptus extract lower enough for root length of Phalaris to exhibit a stimulated response.
Morphological effects in growth may be the secondary manifestation of primary events, caused by variety of more specific effects acting at the cellular or molecular level in the receiver plants. Moreover, the inhibitory compounds might have reduced the uptake of nutrients which ultimately reduced shoot growth (Peng et al., 2004). Khan et al. (2005) also reported that water extracts of Prosopis juliflora and Eucalyptus camaldulensis and bark of Acacia nilotica significantly reduced germination percentage, seedling length and biomass yield of Ipomoea sp., Asphodelus tenuifolius establishment.
Present results revealed that most of the growth parameters of Phalaris were decreased when exposed to different amounts of decompose and water extracts of Eucalyptus leaf. The reduction is more severe by Eucalyptus leaves decompose.
Effects of decompose and aqueous extract of Eucalyptus leaf on antioxidant enzymes activity (catalase, peroxidase, ascorbate peroxidase and poly phenol oxidase) in root and leaf of Phalaris have been showed in Table 4-7.
Poly phenoloxidase and peroxidase activity in root and shoot of Phalaris were not significantly affected by decompose and aqueous extract of Eucalyptus leaf. However, catalase activity in root and leaf of Phalaris was reduced by leaf decompose of Eucalyptus and was increased by high amount of aqueous extract of Eucalyptus leaf. Also, present results indicated that ascorbate peroxidase in Phalaris root was significantly increased by aqueous extract of Eucalyptus leaf.
In general, various types of environmental stresses (including abiotic, xenobiotic and herbicidal) mediate their impact through oxidative stress caused by generation of Reactive Oxygen Species, ROS (Blokhina et al., 2003). ROS, such as singlet oxygen (1O2), hydroxyl radicals (HO•) and hydrogen peroxide (H2O2), are highly reactive and toxic molecules that can cause oxidative damage to membranes, DNA, proteins, photosynthetic pigments and lipids (Apel and Hirt, 2004). Recently, ROS generation and related oxidative stress has been proposed as one of the modes of action of plant growth inhibition by allelochemicals. However, very little is known about the action of allelochemicals/phytotoxins in inducing ROS-mediated oxidative damage. Bais et al. (2003) reported that catechin, a putative phytotoxin, inhibits plant growth due to a severe oxidative burst in root tips, resulting in cell death.
Recently, allelochemicals have been proposed to cause oxidative stress in target tissue and induce an antioxidant mechanism (Li and Hu, 2005; Singh et al., 2006). Activities of the antioxidant enzymes Super Oxide Dismutase (SOD), catalase (CAT), ascorbate peroxidase (APX), peroxidase (PX) were significantly elevated, thereby indicating the enhanced generation of Reactive Oxygen Species (ROS) by allelochemicals (Apel and Hirt, 2004; Niakan et al., 2008). Increased levels of scavenging enzymes indicates their induction as a secondary defense mechanism in response allelopathic compounds.
α-Pinene is one of the major components of volatiles released by a wide range of species throughout the world (May and Ash, 1990; Geron et al., 2001, 2002; Singh et al., 2006; Hartikainen et al., 2009). α-Pinene inhibits seed germination and primary root growth in maize and disrupts energy metabolism by acting as an uncoupler of oxidative phosphorylation and inhibiting the electron transport chain (Abrahim et al., 2003).
To avoid the cellular damage due to ROS generation, plants produce a number of antioxidant enzymes that are induced and provide secondary protection against oxidative stress (Blokhina et al., 2003; Apel and Hirt, 2004). Increased ROS generation due to α-pinene is also indicated by enhanced activities of scavenging enzymes such as SOD, CAT, APX. Activity of SOD in Cassia occidentalis roots increased significantly in response to α-pinene compared with controls (Singh et al., 2006). However, excessive ROS may cause a decrease in SOD and POD activities (Mishra et al., 1993).
Results of this study showed that antioxidant enzymes activity in root and shoot of Phalaris were affected of aqueous extracts and decompose of Eucalyptus leaf. Exposure to decompose and aqueous extract of Eucalyptus leaf induced oxidative stress through the enhanced generation of ROS which was accompanied by change in activation of antioxidant enzymes systems. Thus, we can used of decompose and aqueous extract of Eucalyptus leaf as natural herbicide.
REFERENCES
- Abrahim, D., A.C. Francischini, E.M. Pergo, A.M. Kelmer-Bracht and E.L. Ishii-Iwamoto, 2003. Effects of α-pinene on the mitochondrial respiration of maize seedlings. Plant Physiol. Biochem., 41: 985-991.
CrossRef - Apel, K. and H. Hirt, 2004. Reactive oxygen species: Metabolism, oxidative stress and signal transduction. Annu. Rev. Plant Biol., 55: 373-399.
CrossRefPubMedDirect Link - Arrigoni, O., 1994. Ascorbate system in plant development. J. Bioeng. Biomem., 26: 407-419.
Direct Link - Bais, H.P., R. Vepachedu, S. Gilroy, R.M. Callaway and J.M. Vivanco, 2003. Allelopathy and exotic plant invasion: From molecules and genes to species interactions. Science, 301: 1377-1380.
CrossRefDirect Link - Blokhina, O., E. Virolainen and K.V. Fagerstedt, 2003. Antioxidants, oxidative damage and oxygen deprivation stress: A review. Ann. Bot., 91: 179-194.
CrossRefPubMedDirect Link - Chance, B. and A.C. Maehly, 1955. Assay of catalases and peroxidases. Methods Enzymol., 2: 764-775.
CrossRefDirect Link - Batish, D.R., N. Setia, H.P. Singh and R.K. Kohli, 2004. Phytotoxicity of lemon-scented eucalypt oil and its potential use as a bioherbicide. Crop Protect, 23: 1209-1214.
CrossRef - Duarte, N.F., E.U. Bucek, D. Karam, N. Sa and M.R.M. Scotti, 2006. Mixed field plantation of native and exotic species in semi-arid Brazil. Aust. J. Bot., 54: 755-764.
CrossRefDirect Link - Gechev, T.S. and J. Hille, 2005. Hydrogen peroxide as a signal controlling plant programmed cell death. J. Cell. Biol., 168: 17-20.
CrossRefDirect Link - Geron, C., P. Harley and A. Guenther, 2001. Isoprene emission capacity for US tree species. Atmospheric Environ., 35: 3341-3352.
Direct Link - Geron, C., A. Guenther, J. Greenberg, H.W. Loescher, D. Clark and B. Baker, 2002. Biogenic volatile organic compound emissions from a lowland tropical wet forest in costa rica. Atmospheric Environ., 36: 3793-3802.
Direct Link - Hartikainen, K., A.M. Nerg, M. Kivimaenpaa, S. Kontunen-soppela and M. Maenpaa et al., 2009. Emissions of volatile organic compounds and leaf structural characteristics of European aspen (Populus tremula) grown under elevated ozone and temperature. Tree Physiol., 29: 1163-1173.
CrossRefDirect Link - Khan, M.A., K.B. Marwat, G. Hassan and Z. Hussain, 2005. Bioherbicidal effects of tree extracts on seed germination and growth of crops and weeds. Pak. J. Weed Sci. Res., 11: 89-94.
Direct Link - Kohli, R.K., D.R. Batish and H.P. Singh, 1998. Eucalypt oil for the control of parthenium (Parthenium hysterophorus L.). Crop Prot., 17: 119-122.
CrossRef - Lee, Y.P., S.H. Kim, J.W. Bang, H.S. Lee S.S. Kwak and S.Y. Kown, 2007. Enhanced tolerance to oxidative stress in transgenic tobacco plants expressing three antioxidant enzymes in chloroplasts. Plant Cell. Rep., 26: 591-598.
CrossRefDirect Link - Li, F.M. and H.Y. Hu, 2005. Isolation and characterization of a novel antialgal., allelochemical from Phragmites communis. Applied Environ. Microbial, 71: 6545-6553.
CrossRefDirect Link - Manoranjan, K. and. D.B. Mishram, 1976. Catalase, peroxidase and poly phenol oxidase activities during rice leaf senescence. Plant Biochem. Enz, 57: 315-319.
Direct Link - Mishra, N.P., R.K. Mishra and G.S. Singhal, 1993. Changes in the activities of anti-oxidant enzymes during exposure of intact wheat leaves to strong visible light at different temperatures in the presence of protein synthesis inhibitors. Plant Physiol., 102: 903-910.
Direct Link - Niakan M., M. Tajari and M. Ghorbanli, 2008. Effects of salinity on allelopathic potential of canola (Brassica napus L.). Allelopath J., 21: 329-338.
Direct Link - Pei, Z.M., Y. Murata, G. Benning, S. Thomine and B. Klusener et al., 2000. Calcium channels activated by hydrogen peroxide mediate abscisic acid signalling in guard cells. Nature, 406: 731-734.
CrossRefDirect Link - Peng, S.L., J. Wen and Q.F. Guo, 2004. Mechanism and active variety of allelochemical. Acta Botanica Sinica, 46: 757-766.
Direct Link - Prati, D. and O. Bossdorf, 2004. Allelopathic inhibition of germination by Alliaria petiolata (Brassicaceae). Am. J. Bot., 91: 285-288.
Direct Link - Sasikumar, K., C. Vijayalakshmi and K.T. Parthiban, 2001. Allelophatic effects of four eucalyptus species on redgram (Cajanus cajan L.). J. Trop Agric., 39: 134-138.
Direct Link - Shahid, M., B. Ahmad, R.A. Khattak. G. Hassan and H. Khan, 2006. Response of wheat and its weeds to different allelopathic plant water extracts. Pak. J. Weed Sci. Res., 12: 61-68.
Direct Link - Singh, H.P., D.R. Batish, S. Kaur, K. Arora and R.K. Kohli, 2006. α-Pinene inhibits growth and induces oxidative stress in roots. Ann. Bot., 98: 1261-1269.
CrossRefDirect Link