
F. Mokhberdoran
Department of Agronomy, Faculty of Agriculture, Islamic Azad University of Mashhad Branch, Iran
S.M. Nabavi Kalat
Department of Agronomy, Faculty of Agriculture, Islamic Azad University of Mashhad Branch, Iran
R. Sadrabadi Haghighi
Department of Agronomy, Faculty of Agriculture, Islamic Azad University of Mashhad Branch, Iran
Asian Journal of Plant Sciences
Year: 2009 | Volume: 8 | Issue: 6 | Page No.: 409-416







ABSTRACT
This study was conducted to evaluate seed performance under controlled condition and main aim is to show the effects of NaCl, polyethylene glycol (PEG)-8000, temperature (20, 25 and 30°C) and their interactions on Seed Germination (SG) and Seedling Growth (SDG) of a single rice (Oryza sativa) cultivar which is the name of Kalat. It was obtained from one field harvested in 2008. Seeds were treated with the iso-osmotic concentrations of NaCl or PEG (0, -0.2, -0.4, -0.6 and -0.8 MPa) and exposed to the three temperatures (Namely, 20, 25 and 30°C) for 10 days. There were significant solution types (NaCl or PEG)xtemperaturexosmotic potentials interactions (p<0.05) on the FG, the Germination Rate (GR), the Radicle Length (RL), the Hypocotyl Length (HL), the Seedling Dry Weight (SDW) and the Fresh Weight (FW) indicating that the rice seeds responded differently to salt, drought and temperature changes. The highest values of germination parameters were obtained with no osmotic potential (0 MPa) and increases in osmotic potential either by NaCl or PEG inhibited germination indices. The results indicated that the reduction of germination characteristics was much greater for the PEG induced stress compared to the NaCl induced stress at all osmotic and all thermal levels. In addition, the rice seeds exhibited strong effect of change in temperature on germination characteristics. The deleterious effect of NaCl and PEG was more pronounced at 20 and 30°C compared to 25°C. The examined rice cultivar was more tolerant of NaCl salinity and water stress stimulated by the PEG in the germination stage than in the seedling developmental phase.
PDF Abstract XML References Citation
How to cite this article
F. Mokhberdoran, S.M. Nabavi Kalat and R. Sadrabadi Haghighi, 2009. Effect of Temperature, Iso-Osmotic Concentrations of NaCl and PEG Agents on Germination and Some Seedling Growth Yield Components in Rice (Oryza sativa L.). Asian Journal of Plant Sciences, 8: 409-416.
DOI: 10.3923/ajps.2009.409.416
URL: https://scialert.net/abstract/?doi=ajps.2009.409.416
DOI: 10.3923/ajps.2009.409.416
URL: https://scialert.net/abstract/?doi=ajps.2009.409.416
INTRODUCTION
Plants are constantly confronted with various biotic and abiotic stress factors such as low or high temperature, salt, drought, flooding, heat, oxidative stress and heavy metal toxicity (Mahajan and Tuteja, 2005; Achuo et al., 2006; Jaleel et al., 2007). Climate change stemmed from anthropogenic perturbations has led to accelerated occurrence of some stress factors. Among the stages of the plant life cycle, seed germination and seedling emergence and establishment are key processes in the survival and growth of plants (Hadas, 2004). Seeds and seedlings of plants are more prone to stress than adults due to being exposed to higher extreme environmental fluctuations at or near the soil surface (Dodd and Donovan, 1999). Out of the stress factors, drought and salinity are major abiotic constraints that pose serious threat to crop production throughout the world (Zhu, 2001; Hu and Schmidhalter, 2005; Soltani et al., 2006; Yang et al., 2007). It has been estimated that over 26 and 20% of cultivated lands worldwide are affected by drought and salinity stresses (Flowers and Flowers, 2005; Cicek and Cakırlar, 2008). Moreover, salinity is responsible for degradation of 2 million hectares of world agricultural lands every year (Cicek and Cakırlar, 2008). Salt stress is more acute in arid and semiarid regions (Sadat Noori and McNeilly, 2000; Al-Karaki, 2001; Villa-Castorena et al., 2003) where are typically featured by high evaporation rate, low rainfall, high water table and salty irrigation water.
Increasing salinity can influence seed germination by creating osmotic effect, which impedes seed water uptake or through toxic effects of specific ions such as Na+, Cl¯ and SO42¯ (Al-Karaki, 2001; Tobe et al., 2003; Kaya et al., 2006; Shaikh et al., 2007). Resulting in both retardation or delayed seed germination (Song et al., 2006; Guan et al., 2009). Low water potential in soil medium caused by either soil salinity or water deficit, leading to late and inadequate germination and associated failure of stand establishment adversely affects crop productivity (Willenborg et al., 2005). NaCl as ionic stress agent and PEG as non-ionic stress agent have been widely used to study agronomic crops, halophytes and wild species response to salt and drought stresses during germination stage tomato (Solanum lycopersicum L.) mutants (Fellner and Sawhney, 2001); durum wheat (Triticum durum Desf.) (Almansouri et al., 2001) phaseolus (Jeannette et al., 2002); sorghum (Sorghum bicolor L.) (Kader and Jutzi, 2002); caldén (Prosopis caldenia Burk) (De Villalobos et al., 2002); oakleaf goosefoot (Chenopodium glaucum L.) (Duan et al., 2004); argentine screwbean (Prosopis strombulifera) (Sosa et al., 2005); oat (Avena fatua L.) (Willenborg et al., 2005); cowpea (Vigna unguiculata L.) (Murillo-Amador et al., 2006); sugar beet (Beta vulgaris L.) (Jafarzadeh and Aliasgharzad, 2007); rice (Oryza sativa L.) (Aqeel Ahmad et al., 2007). The PEG-induced inhibition of germination has been attributed to osmotic stress (Dodd and Donovan, 1999; Sidari et al., 2008). Temperature plays a crucial role in many biological and physiological processes of plants (Al-Ahmadi and Kafi, 2007; Berti and Johnson, 2008). The temperature changes have major impact on a number of processes which regulate seed germinability, including membrane permeability and the activity of membrane-bound as well as cytosolic enzymes (Tlig et al., 2008) and its interaction with the variable soil water content in the surface layers of the soil, where crop seeds are sown, is critically important in terms of germination onset and rate (Khan et al., 2000; Finch-Savage et al., 2001). Salinity-temperature interaction, in particular, determines seed germination pattern in many salt-affected environments (Khan and Gulzar, 2003; Al-Khateeb, 2006; Song et al., 2006). Rice (Oryza sativa L.) is the second main stable food in Iran following common wheat (Triticum aestivum L.). However, its production has been drastically decreased due to salinity and drought, especially in those areas that rice is directly sown into puddled and leveled soil. Rice (Oryza sativa L.) is considered as a salt sensitive crop compared to other cereals but its sensitivity to salinity varies greatly with developmental stage (Zeng and Shannon, 2000) and among cultivars (Quijano-Guerta and Kirk, 2002). Kalat cultivar used in this study is locally cultivated in the Northeastern parts of Iran which is currently experiencing difficulties with the soil salinization and frequent droughts. Therefore, it is necessary to evaluate the rice seeds response to such abiotic stresses.
The aim of this study is to investigate the effects of osmotic stress generated by NaCl or PEG and combined effects of these stress factors with temperature on germination characteristics and SDG of rice (Oryza sativa L.) cultivar during seed germination.
MATERIALS AND METHODS
This study was carried out at the Faculty of Agriculture, Islamic Azad University of Mashhad, Iran from May till August 2008. Seeds of Kalat cultivar were used in this study. It was provided from one field harvested in 2008 at Kalat region, Iran. Germination tests and seedling evaluation were performed according to ISTA guidelines (Kaya et al., 2006). Seeds were surfaced sterilized in 1% sodium hypochlorite solution for 1 min, then were rinsed two times for one minute with sterile distilled water (Alam et al., 2002). The experiment was a completely randomized design with a factorial arrangement consisting of two osmotic solutions (PEG or NaCl), five osmotic potentials (0, -0.2, -0.4, -0.6 and -0.8 MPa) and three constant temperature degrees (20, 25 and 30°C) with three replications. Osmotic potential levels for the PEG-8000 were obtained using the method described by Michel and Kaufmann (1973) and for the NaCl were created based on the van't Hoff's equation (Ben-Gal et al., 2009):
Π = iMRT |
where, Π is osmotic potential (MPa), i is the van't Hoff factor (moles of particle in solution per moles of dissolved solute), M is molarity of the solute (mol L-1), R is the universal gas law constant (8.314x10-6 m-3 MPa mol-1 K-1) and T is temperature (298K).
Twenty rice seeds were placed and germinated into sterilized petri dishes (Π= 9 cm) on Whatman No.1 filter paper. Each petri dish was moistened with 10 mL of distilled water (control) or the respective test solutions (Song et al., 2006). The Petri dishes were covered with lids and placed in incubators in the dark at the 20, 25 and 30°C (Guan et al., 2009). The solutions (the NaCl and PEG-8000) were changed every other day to keep the primary water potential. Germination counting was done at 24 h intervals for 10 days (Kaya et al., 2008). Seeds were considered as germinated when radicle had protruded 2 mm through the seed coat. After 10 days of incubation, FG (%), GR (seeds day-1), RL (mm), HL (mm), seedling FW (g seedling-1) and DW (g seedling-1) were determined (Murillo-Amador et al., 2002). Rate of germination (seeds day-1) was estimated using Maguire`s equation (Pezzani and Montana, 2006):
Rs = ∑ Si/Di |
where, Rs is GR (the number of germinated seeds per day), Si is the number of germinated seeds at each counting and Di is number of days until the nth count. Dry weights of seedlings (g seedling-1) were measured after drying samples at 70°C for 48 h in an oven (Okcu et al., 2005).
After completion of the germination tests non-germinated seeds were transferred to distilled water at the initial temperatures for further 5 days to determine the toxic effects of the solutions on germination (Khan et al., 2001). The obtained data were analyzed using the MSTAT-C statistical software and the differences between means were compared by Duncan’s multiple range at probability level of 5% (Jafarzadeh and Aliasgharzad, 2007).
RESULTS AND DISCUSSION
Germination: Results of the analysis of variance (ANOVA) for the FG and GR showed that there were significant differences between all treatments and their interactions, except solutionsxtemperature interaction for the FG (Table 1). At 20°C under -0.8 MPa, NaCl and PEG treatments, the FG was declined to 49.9 and 90% of control, respectively, while at 25°C under -0.8 MPa NaCl and PEG concentration 88 and 50% of seeds germinated, respectively. The decrease in the FG under the highest level of NaCl was 25% of control treatment at 30°C, while this value under PEG treatment was 85% of control (Table 2). Transfer of non- germinated seeds from PEG solution to the distilled water resulted in 100% germination recovery at all temperatures and osmotic potential levels. But there was little germination recovery for the NaCl solution, particularly at the 20 and 30°C under the moderate and highest salinity levels (data not shown). At the 25°C and osmotic potential of -0.8 MPa, the GR of NaCl-treated seeds was 3.9 seeds day-1, while the GR of PEG-treated seeds was 1.2 seeds day-1. At 30 and 20°C under -0.8 Mpa NaCl the GR was 2.2 and 0.6 seed day-1, respectively. For -0.8 MPa, PEG treatment on the GR was 0.2 and 0.1 seeds day-1 at the 30 and 20°C, respectively (Table 3).
Seedling growth: The ANOVA for the RL and HL parameters showed that all factors and all interactions were significant (Table 4). The reduction in the RL at the 25°C under the moderate and highest osmotic potentials of NaCl (-0.4 and -0.8 MPa) varied between 18 and 32% of control treatment, while at the same temperature and comparable osmotic potentials in the PEG the reduction varied between 50 and 75% of control treatment, respectively. At the 20 and 30°C under the -0.4 MPa PEG, the RL was decreased by approximately 62% when it was compared with their respective control treatments. Also at the 20 and 30°C under -0.4 MPa, NaCl the decrease of the RL varied between 39 and 42% of their respective control treatments (Table 5). The NaCl treatment at the 25°C under osmotic potentials of the -0.4 and -0.8 MPa, declined the HL to 28.1 and 54.3% of control treatment, respectively, while the PEG treatment at similar temperature and comparable osmotic potentials declined the HL to 70.2 and 85.1% of control. -0.4 MPa, NaCl stress at the 20 and 30°C decreased the HL by about 47.1 and 43.4% of their respective control treatments, while -0.4 MPa, the PEG stress at the 20 and 30°C caused a reduction of 77.5 and 84% in the HL when it was compared with their respective control treatments. Moreover, no the HL was recorded at the 20°C above osmotic potential of -0.4MPa, the PEG and at the 30°C at the -0.8MPa, the PEG solution (Table 6).
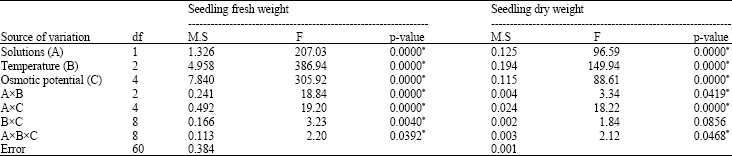
Seedling weight (g seedling-1): The ANOVA for the seedling FW and DW showed significant differences between all factors and their interactions except temperaturexosmotic potential for the SDW (Table 7). The decrease in the seedling FW at the 25 and 30°C under the highest NaCl level ranged between 39 and 58% of their respective control treatments, while for the PEG stress at the 20 and 30°C under the highest concentrations the decrease was about 91% of their respective control treatments. At the 20°C, a 52 and 67% decrease in the seedling FW was observed in the moderate osmotic potentials of the NaCl and the PEG (-0.4 MPa), respectively (Table 8).
| Table 1: | Variance analysis results of the seed germination and germination rate in used rice cultivar |
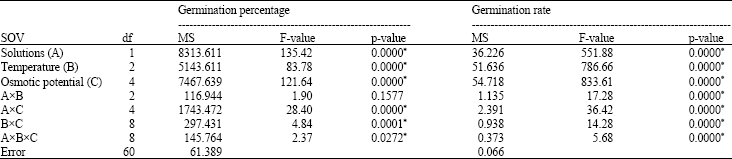 | |
| *Significant at p-value of 0.05 | |
| Table 2: | Effects of NaCl, PEG and temperature on germination percentage (%) |
 | |
| Means with similar superscript letter(s) are not significantly different (p<0.05) | |
| Table 3: | Effects of NaCl, PEG and temperature on germination rate (seeds day-1) |
 | |
| Means with similar superscript letter(s) are not significantly different (p<0.05) | |
| Table 4: | Variance analysis results of the radicle and hypocotyl lengths in used rice cultivar |
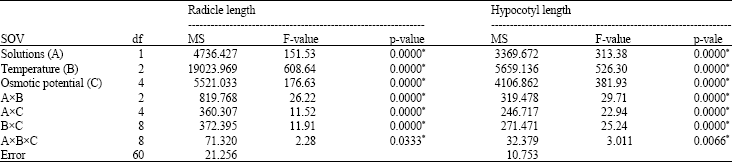 | |
| *Significant at p-value of 0.05 | |
| Table 5: | Effects of NaCl, PEG and temperature on radicle length (mm) |
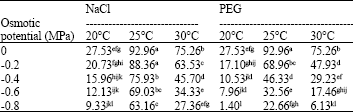 | |
| Means with similar superscript letter(s) are not significantly different (p<0.05) | |
| Table 6: | Effects of NaCl, PEG and temperature on hypocotyl length (mm) |
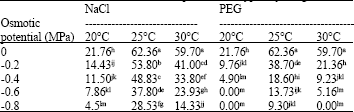 | |
| Means with similar superscript letter(s) are not significantly different (p<0.05) | |
At the 25 and 30°C the highest osmotic potential of the NaCl resulted in a reduction in the SDW by only about 19% when it was compared with their respective control treatments, while the decrease in the SDW at the 25 and 30°C under the highest PEG level fluctuated between 42 and 83% of control treatment. At the 20°C, the SDW exhibited a decrease of 40% at -0.8 MPa, NaCl, while the PEG at similar temperature and comparable osmotic potential diminished the SDW by 85% of control (Table 9).
Overall, the application of the NaCl and PEG at different thermal levels decreased germination characteristics. Considerable variation in the rice SG response to temperature alterations and solute type was observed. Temperature and osmotic moist stress interacted with one another in the germination of rice seed. Germination indices were less affected at the 25°C, particularly under salinity stress and any decrease or increase from this level was associated with accelerated effects of two stress factors. This indicates that, moving away from optimal to over-optimal or sub-optimal germination temperature significantly increases of the rice sensitivity to salt or drought stress. This observation is consistent with the studies on a range of plant species such as red swampfire (Salicornia rubra) (Khan et al., 2000); carrot (Daucus carota L.) (Finch-Savage et al., 2001); kochia (Kochia scoparia) (Khan et al., 2001); globe artichoke (Cynara cardunculus) (Mauromicale and Licandro, 2002); canola (Brassica napus L.) (Willenborg et al., 2004); mesquite (Prosopis juliflora) (El-Keblawy and Al-Rawai, 2005); desert grass (Panicum turgidum Forssk) (Al-Khateeb, 2006); alfalfa (Medicago ruthenica L.) (Guan et al., 2009) where the injury effects of NaCl or PEG are generally less severe at optimum temperature. It can be proposed that enzymatic functions of seeds are more sensitive to stressful condition when temperature is not convenient. Furthermore, higher temperature could promote ions activities especially Na+ and Cl¯ which results in ions fluxes through cell membrane and the increased risk of toxicity to seed germination process. High osmoticum levels such as the PEG agent, high salinity and low temperature impair water absorption and transport in seeds (Fellner and Sawhney, 2001).
| Table 7: | Variance analysis results of the seedling fresh and dry weights in used rice cultivar |
 | |
| *Significant at p-value of 0.05 | |
| Table 8: | Effects of NaCl, PEG and temperature on seedling fresh weight (g seedling-1) |
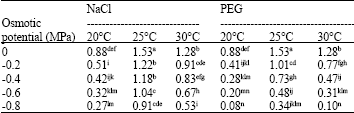 | |
| Means with similar superscript letter(s) are not significantly different (p<0.05) | |
| Table 9: | Effects of NaCl, PEG and temperature on seedling dry weight (g seedling-1) |
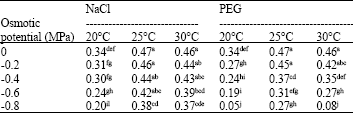 | |
| Means with similar superscript letter(s) are not significantly different (p<0.05) | |
The inhibition of GR was stronger at different levels of the NaCl and PEG solutions compared to the FG percentage. This result corroborates other studies showing that osmotic stress primarily reduces rate of germination rather than germination percentage (Alam et al., 2002; Atak et al., 2006; Kaya et al., 2008). Velocity of the reduction in the values of germination and seedling growth of rice was more profound under the PEG solute than the NaCl, irrespective of temperature changes. Similar results were found in durum wheat (Triticum durum Desf.) by Almansouri et al. (2001) in cowpea (Vigna unguiculata L.) by Murillo-Amador et al. (2002) in saxaul (Haloxylon ammodendron) by Tobe et al. (2004) in pea (Pisum sativum L.) by Okcu et al. (2005) and in sunflower (Helianthus annuus L.) by Kaya et al. (2006). Tobe et al. (2004) and Kaya et al. (2006) mentioned that the uptake of external ions (Na+, Cl¯) by the seed, maintaining a water potential gradient which allows water uptake during seed germination under NaCl stress. Salt and drought stress inhibited the growth of hypocotyl more than radicle. This observation is supported by other studies on corn (Zea mays L.) (Parmar and Moore, 1968); rice (Oryza sativa L.) (Alam et al., 2002); Phaseolus (Jennertte et al., 2002); triticale (x Triticosecale Wittmack) (Atak et al., 2006). Parmar and Moore (1968) attributed less adverse effects of the NaCl and PEG on radicle growth to the greater dependence of the radicle on the seed reserve storage and less dependence on water uptake. However, these findings are inconsistent with those of Jamil et al. (2007) who, observed that the reduction in RL of radish seeds by increased salinity was more prominent compared to the HL. It appears that in most crop seeds such as corn (Zea mays L.), pea (Pisum sativum L.) and rice (Oryza sativa L.) HL is more adversely affected than RL by both toxic and osmotic effects as compared to other plants.
The results of this study showed a greater inhibition of HL and RL in comparison to germination phase by decreasing osmotic potential with NaCl and PEG induced stress. This is in conformity with findings from Khajeh Hosseini et al. (2002) in soybean; Okcu et al. (2005) in pea (Pisum sativum L.) (2005) and Kaya et al. (2008) in chickpea (Cicer arietinum L.) who observed that NaCl or PEG had greater inhibitory effects on seedling development than germination. Khajeh Hosseini et al. (2002) and Alam et al. (2002) suggested that cell division, which is a post-germination phenomenon responsible for seedling elongation and development, is more sensitive to the NaCl or PEG compared to cell expansion, which drives germination. The seedling FW (g seedling-1) and DW (g seedling-1) were diminished as a result of decrease in seedling growth under osmotic stress. However, the DWs were less affected than fresh weights in the NaCl and PEG. Murillo-Amador et al. (2002) in cowpea (Vigna unguiculata L.) reported that the SDW decreased with lower magnitude compared to the seedling FW under varying levels of NaCl and PEG.
In conclusion, the results of this study revealed that the rice (Oryza sativa L.) germination was very responsive to temperature alterations and temperature was main source of variation of the SDG. This is important in terms of predicting germination time under variable conditions of the temperature and soil moisture (Finch-Savage et al., 2001) which would be helpful for farmers in taking management decisions. On the one hand, below temperature values of the 20°C in combination with osmotic stress greatly reduced the FG percentage and the GR as well as the SDG. On the other hand, increasing temperature from 25 to 30°C accentuated the deleterious effects of the osmotic stress on the measured traits. Best germination value was obtained at 25°C. Moreover, used rice cultivar in this study was better able to cope with salinity stress during the germination and SDG stages when the optimal temperature of the 25°C prevailed. On the other hand, seed germination was found to be less sensitive to the NaCl and PEG agents than subsequent stages. Furthermore, investigated all germination parameters responses to the PEG treatment appeared to have been osmotically induced, rather than toxic effects of the NaCl.
REFERENCES
- Al-Ahmadi, M.J. and M. Kafi, 2007. Cardinal temperatures for germination of Kochia scoparia (L.). J. Arid Environ., 68: 308-314.
CrossRef - Alam, M.Z., T. Stuchbury and R.E.L. Naylor, 2002. Effect of NaCl and PEG induced osmotic potentials on germination and early seedling growth of rice cultivars differing in salt tolerance. Pak. J. Biol. Sci., 5: 1207-1210.
CrossRefDirect Link - Al-Karaki, G.N., 2001. Germination, sodium and potassium concentrations of barley seeds as influenced by salinity. J. Plant Nutr., 24: 511-522.
CrossRefDirect Link - Al-Khateeb, S.A., 2006. Effect of salinity and temperature on germination, growth and ion relations of Panicum turgidum forssk. Bioresour. Technol., 97: 292-298.
CrossRefDirect Link - Almansouri, M., J.M. Kinet and S. Lutts, 2001. Effect of salt and osmotic stresses on germination in durum wheat (Triticum durum Desf.). Plant Soil, 231: 243-254.
CrossRefDirect Link - Ahmad, M.S.A., F. Javed and M. Ashraf, 2007. Iso-osmotic effect of NaCl and PEG on growth, cations and free proline accumulation in callus tissue of two indica rice (Oryza sativa L.) genotypes. Plant Growth Regul., 53: 53-63.
CrossRefDirect Link - Atak, M., M.D. Kaya, G. Kaya, Y. Cikili and C.Y. Ciftci, 2006. Effects of NaCl on the germination, seedling growth and water uptake of triticale. Turk. J. Agric. For., 30: 39-47.
Direct Link - Ben-Gal, A., H. Borochov-Neori, U. Yermiyahu and U. Shani, 2009. Is osmotic potential a more appropriate property than electrical conductivity for evaluating whole-plant response to salinity?. Environ. Exp. Bot., 65: 232-237.
CrossRefDirect Link - Berti, M.T. and B.L. Johnson, 2008. Seed germination response of cuphea to temperature. Ind. Crops Prod., 27: 17-21.
CrossRefDirect Link - Cicek, N. and H. Cakırlar, 2008. Effects of salt stress on some physiological and photosynthetic parameters at three different temperatures in six soya bean (Glycine max L. Merr.). J. Agron. Crop Sci., 194: 34-46.
CrossRefDirect Link - De Villalobos, A.E., D.V. Pelaezwz, R.M. Boowz, M.D. Mayorz and O.R. Elia, 2002. Effect of high temperatures on seed germination of Prosopis caldenia Burk. J. Arid Environ., 52: 371-378.
CrossRef - Dodd, G.L. and L.A. Donovan, 1999. Water potential and ionic effects on germination and seedling growth of two cold desert shrubs. Am. J. Bot., 86: 1146-1153.
PubMedDirect Link - Duan, D., X. Liu, M.A. Khan and B. Gul, 2004. Effects of salt and water stress on the germination of Chenopodium glaucum L. seed. Pak. J. Bot., 36: 793-800.
Direct Link - El-Keblawy, A. and A. Al-Rawai, 2005. Effects of salinity, temperature and light on germination of invasive Prosopis juliflora (Sw.) D.C. J. Arid Environ., 61: 555-565.
CrossRefDirect Link - Fellner, M. and V.K. Sawhney, 2001. Seed germination in a tomato male-sterile mutant is resistant to osmotic, salt and low-temperature stresses. Theor. Appl. Genet., 102: 215-221.
CrossRefDirect Link - Finch-Savage, W.E., K. Phelps, J.R.A. Steckel, W.R. Whalley and H.R. Rowse, 2001. Seed reserve dependent growth responses to temperature and water potential in carrot (Daucus carota L.). J. Exp. Bot., 52: 2187-2197.
PubMedDirect Link - Flowers, T.J. and S.A. Flowers, 2005. Why does salinity pose such a difficult problem for plant breeders? Agric. Water Manage., 78: 15-24.
CrossRef - Guan, B., D. Zhou, H. Zhang, Y. Tian, W. Japhet and P. Wang, 2009. Germination responses of Medicago ruthenica seeds to salinity, alkalinity and temperature. J. Arid Environ., 73: 135-138.
CrossRefDirect Link - Hu, Y. and U. Schmidhalter, 2005. Drought and salinity: A comparison of their effects on mineral nutrition of plants. J. Plant Nutr. Soil Sci., 168: 541-549.
CrossRefDirect Link - Jafarzadeh, A.A. and N. Aliasgharzad, 2007. Salinity and salt composition effects on seed germination and root length of four sugar beet cultivars. Biologia, 62: 562-564.
CrossRef - Jaleel, C.A., P. Manivannan, B. Sankar, A. Kishorekumar, R. Gopi, R. Somasundaram and R. Panneerselvam, 2007. Water deficit stress mitigation by calcium chloride in Catharanthus roseus: Effects on oxidative stress, proline metabolism and indole alkaloid accumulation. Colloids Surf. B: Biointerfaces, 60: 110-116.
CrossRefPubMedDirect Link - Jamil, M., Shafiq ur Rehman, K.J. Lee, J.M. Kim, H.S. Kim and E.U. Rha, 2007. Salinity reduced growth PS2 photochemistry and chlorophyll content in radish. Sci. Agric., 64: 111-118.
CrossRefDirect Link - Kader, M.A. and S.C. Jutzi, 2002. Temperature, osmotic pressure and seed treatments influence imbibition rates in sorghum seeds. J. Agron. Crop Sci., 188: 286-290.
CrossRefDirect Link - Kaya, M.D., G. Okcu, M. Atak, Y. Cikili and O. Kolsarici, 2006. Seed treatments to overcome salt and drought stress during germination in sunflower (Helianthus annuus L.). Eur. J. Agron., 24: 291-295.
CrossRefDirect Link - Kaya, M., G. Kaya, M.D. Kaya, M. Atak, S. Saglam, K.M. Khawar and C.Y. Ciftci, 2008. Interaction between seed size and NaCl on germination and early seedling growth of some Turkish cultivars of chickpea (Cicer arietinum L.). J. Zhejiang Univ. Sci. B, 9: 371-377.
CrossRefDirect Link - Hosseini, M.K., A.A. Powell and I.J. Bingham, 2002. Comparison of the seed germination and early seedling growth of soybean in saline conditions. Seed Sci. Res., 12: 165-172.
CrossRefDirect Link - Khan, M.A., B. Gul and D.J. Weber, 2000. Germination response of Salicornia rubra to temperature and salinity. J. Arid Environ., 45: 207-214.
CrossRef - Khan, M.A., B. Gul and D.J. Weber, 2001. Influence of salinity and temperature on the germination of Kochia scoparia. Wetlands Ecol. Manage., 9: 483-489.
CrossRefDirect Link - Khan, M.A. and S. Gulzar, 2003. Light, salinity and temperature effects on the seed germination of perennial grasses. Am. J. Bot., 90: 131-134.
Direct Link - Mahajan, S. and N. Tuteja, 2005. Cold, salinity and drought stresses: An overview. Arch. Biochem. Biophys., 444: 139-158.
CrossRefPubMedDirect Link - Mauromicale, G. and P. Licandro, 2002. Salinity and Temperature effects on germination, emergence and seedling growth of globe artichoke. Agronomie, 22: 443-450.
CrossRefDirect Link - Michel, B.E. and M.R. Kaufmann, 1973. The osmotic potential of polyethylene glycol 6000. Plant Physiol., 51: 914-916.
CrossRefPubMedDirect Link - Murillo-Amador, B., R. Lopez-Aguilar, C. Kaya, J. Larrinaga-Mayoral and A. Flores-Hernandez, 2002. Comparative effects of NaCl and polyethylene glycol on germination, emergence and seedling growth of cowpea. J. Agron. Crop Sci., 188: 235-247.
CrossRefDirect Link - Murillo-Amador, B., E. Troyo-Dieguez, J.L. Garcia-Hernandez, R. Lopez-Aguilar and N.Y. Avila-Serrano et al., 2006. Effect of NaCl salinity in the genotypic variation of cowpea (Vigna unguiculata) during early vegetative growth. Scientia Hortic., 108: 432-441.
CrossRefDirect Link - Okcu, G., M.D. Kaya and M. Atak, 2005. Effects of salt and drought stresses on germination and seedling growth of pea (Pisum sativum L.). Turk. J. Agric. For., 20: 237-242.
Direct Link - Parmar, M.T. and R.P. Moore, 1968. Carbowax 6000, mannitol and sodium chloride for simulating drought conditions in germination studies of corn (Zea mays L.) of strong and weak vigor. Agron. J., 60: 192-195.
Direct Link - Pezzani, F. and C. Montana, 2006. Inter-and intra-specific variation in the germination response to light quality and scarification in grasses growing in two-phase mosaics of the chihuahuan desert. Ann. Bot., 97: 1063-1071.
CrossRef - Quijano-Guerta, C. and G.J.D. Kirk, 2002. Tolerance of rice germplasm to salinity and other soil chemical stresses in tidal wetlands. Field Crop Res., 76: 111-121.
CrossRef - Sadat Noori, S.A. and T. McNeilly, 2000. Assessment of variability in salt tolerance based on seedling growth in Triticum durum desf. Genet. Resour. Crop Evol., 47: 285-291.
CrossRefDirect Link - Shaikh, F., B. Gul, W.Q. Li, X.J. Liu and M.A. Khan, 2007. Effect of calcium and light on the germination of Urochondra setulosa under different salts. J. Zhejiang Univ. Sci. B, 8: 20-26.
CrossRefPubMedDirect Link - Sidari, M., C. Mallamaci and A. Muscolo, 2008. Drought, salinity and heat differently affect seed germination of Pinus pinea. J. Forest Res., 13: 326-330.
CrossRefDirect Link - Soltani, A., M. Gholipoor and E. Zeinali, 2006. Seed reserve utilization and seedling growth of wheat as affected by drought and salinity. Environ. Exp. Bot., 55: 195-200.
Direct Link - Song, J., G. Feng and F. Zhang, 2006. Salinity and temperature effects on germination for three salt-resistant euhalophytes, Halostachys caspica, Kalidium foliatum and Halocnemum strobilaceum. Plant Soil, 279: 201-207.
CrossRef - Sosa, L., A. Llanes, H. Reinoso, M. Reginato and V. Luna, 2005. Osmotic and specific ion effects on the germination of Prosopis strombulifera. Ann. Bot., 96: 261-267.
CrossRef - Tlig, T., M. Gorai and M. Neffati, 2008. Germination responses of Diplotaxis harra to temperature and salinity. Flora-Morphol. Distrib. Funct. Ecol. Plants, 203: 421-428.
CrossRef - Tobe, K., L. Zhang and K. Omasa, 2003. Alleviatory effects of calcium on the toxicity of sodium, potassium and magnesium chlorides to seed germination in three halophytes. Seed Sci. Res., 13: 47-54.
CrossRef - Tobe, K., L. Xiaoming and K. Omasa, 2004. Effects of five different salts on seed germination and seedling growth of Haloxylon ammodendron (Chenopodiaceae). Seed Sci. Res., 14: 345-353.
CrossRef - Villa-Castorena, M., A.L. Ulery, A. Catalan-Valencia and M.D. Remmenga, 2003. Salinity and nitrogen rate effects on the growth and yield of chile pepper plants. Soil Sci. Soc. Am. J., 67: 1781-1789.
Direct Link - Willenborg, C.J., R.H. Gulden, E.N. Johnson and S.J. Shirtliffe, 2004. Germination characteristics of polymer-coated canola (Brassica napus L.) seeds subjected to moisture stress at different temperatures. Agron. J., 96: 786-791.
CrossRefDirect Link - Willenborg, C.J., J.C. Wildeman, A.K. Miller, B.G. Rossnagel and S.J. Shirtliffe, 2005. Oat germination characteristics differ among genotypes, seed sizes and osmotic potential. Crop Sci., 45: 2023-2029.
CrossRefDirect Link - Yang, J., J. Zhang, K. Liu, Z. Wang and L. Liu, 2007. Involvement of polyamines in the drought resistance of rice. J. Exp. Bot., 58: 1545-1555.
CrossRef - Zeng, L. and M.C. Shannon, 2000. Salinity effects on seedling growth and yield components of rice. Crop Sci., 40: 996-1003.
CrossRefDirect Link - Achuo, E.A., E. Prinsen and M. Hofte, 2006. Influence of drought, salt stress and abscisic acid on the resistance of tomato to Botrytis cinerea and Oidium eolycopersic. Plant Pathol., 55: 178-186.
CrossRefDirect Link