
A. Javaid
Faculty of Agriculture, Kagawa University, Ikenobe, Miki Cho, Kita Gun, Kagawa 761-0795, Japan
S. Ikeda
Faculty of Agriculture, Kagawa University, Ikenobe, Miki Cho, Kita Gun, Kagawa 761-0795, Japan
I. Kataoka
Faculty of Agriculture, Kagawa University, Ikenobe, Miki Cho, Kita Gun, Kagawa 761-0795, Japan
S. Taketa
Research Institute for Bioresources, Okayama University, Kurashiki 710-0046, Japan
Asian Journal of Plant Sciences
Year: 2009 | Volume: 8 | Issue: 6 | Page No.: 400-408







ABSTRACT
Short rachilla hair in barley is controlled by a recessive gene srh that is located on the long arm of barley chromosome 5H. It shows a skewed geographical distribution toward the Occidental regions of barley cultivation. This morphological character is an attractive trait to study the phylogeny of barley. In a previous study, srh was mapped between two markers at a distance of 1.6 and 0.5 cm in the proximal and distal side, respectively. An integrated map was constructed during this experiment using the data from two F2 populations and 320 homozygous recessive short rachilla hair plants from another population. The CAPS marker k06288KU and SSR marker Bmag0223 are found to be flanking short rachilla hair locus srh at a genetic distance of 1.6 and 4.0 cm, respectively. World barley germplasm collections of 192 accessions differing in rachilla type were surveyed using k06288KU and Bmag0223. SSR marker Bmag0223 showed a better association with srh. In Europe, North Africa and Ethiopia, the frequency of Bmag0223 allele A was high regardless of rachilla type, but in East Asia allele A was closely associated with short rachilla. On the basis of marker association and genetic diversity, the origin of short rachilla hair character in barley is discussed.
PDF Abstract XML References Citation
How to cite this article
A. Javaid, S. Ikeda, I. Kataoka and S. Taketa, 2009. Fine Mapping of Short Rachilla Hair Gene (srh) in Barley and an Association Study using Flanking Molecular Markers and World Germplasms. Asian Journal of Plant Sciences, 8: 400-408.
DOI: 10.3923/ajps.2009.400.408
URL: https://scialert.net/abstract/?doi=ajps.2009.400.408
DOI: 10.3923/ajps.2009.400.408
URL: https://scialert.net/abstract/?doi=ajps.2009.400.408
INTRODUCTION
Barley (Hordeum vulgare L.) is the fourth most important cereal crop in the world after corn, rice and wheat. It was one of the first crops domesticated and has a significant role in the history of agriculture, especially, during the development of Neolithic culture (Salamini et al., 2002). Barley can be grown in various environmental conditions and has developed a diversity of easily classifiable hereditary morphological traits during evolution (Poehlman, 1987). Linkage maps for these morphological characters are developed by the conventional mapping procedures and molecular techniques and a large number of studies have been directed at research on the evolution and genetic diversity of morphological traits in barley (Bothmer et al., 2003). Barley geneticists are doing intensive research on morphological traits that are important to study the evolution and genetic diversity. Takahashi (1955) provided evidence of genetic differentiation between two distinct groups in world barley; an oriental group and an occidental group, based on his genetic analyses of rachis brittleness in hybrids. He classified barleys grown in China, Southern Japan and South Korea as the oriental group and barleys from the rest of the world as the Occidental group. Komatsuda and Mano (2002) conducted molecular mapping of non-brittle rachis. Komatsuda et al. (2004) reported that non-brittle rachis of Occidental barley lines is controlled by btr1 on chromosome 3H, while, non-brittle rachis of Oriental barley lines is controlled by a major gene btr2 on chromosome 3H and two quantitative trait loci on chromosomes 5HL and 7H. This result suggests at least two independent mutations in the formation of non-brittle rachises in barley. Tanno et al. (2002) studied the origin of six-rowed domesticated barley using a DNA marker cMWG699 closely linked to the vrs1 locus. Kikuchi et al. (2003) developed a fine map around the nud locus controlling naked caryopsis on the long arm of chromosome 7H using High Efficiency Genome Scanning (HEGS) electrophoresis combined with Amplified Fragment Length Polymorphism (AFLP). Taketa et al. (2004) studied the origin of naked barley based on molecular variation of the SCAR marker sKT7 tightly linked to the nud locus. It was concluded that naked barley has a monophyletic origin, probably in Southwestern Iran. The geographical distribution suggested migration routes of naked domesticated barley in Central and Eastern Asia.
Rachilla is the diminutive axis of spikelet in grasses that bears the florets. The rachilla in barley is markedly reduced to a tiny rod-shaped organ. Genetic variation exists in rachilla hair length (Fig. 1). Long rachilla hair is normally observed and short rachilla hair is conditioned by a single recessive gene; thus a simply inherited rachilla hair trait has been used to assess genetic diversity in barley by plant genetic resources scientists and for testing the distinctiveness and uniformity of varieties by barley breeders (Parzies et al., 2000). Although, the adaptive importance of rachilla hair type is not known, it is among the best taxonomic characteristics in barley (Takahashi, 1955; Kebebew et al., 2001). Characterization of barley germplasm from different countries at International Center for Agricultural Research in the Dry Areas (ICARDA) for morphological characters showed the highest diversity for rachilla hair, growth class and growth habit. Short rachilla hair gene srh is located on the long arm of chromosome 5H, about 26.8 cm proximal from the smooth awn gene raw1 (Kleinhofs et al., 1993; Franckowiak, 1997; Lundqvist et al., 1997). It is worth to mention that whereas almost all the varieties of the oriental region have long rachilla hair, varieties with short rachilla hair are frequently found in the occidental region (Takahashi, 1955). Costa et al. (2001) constructed a linkage map of barley using morphological and molecular markers in oregon wolfe barley and reported that Restriction Fragment Length Polymorphism (RFLP) marker MWG522 and Simple Sequence Repeat (SSR) marker Bmag0223 on chromosome 5H are closely linked to short rachilla hair gene srh. Kraakman et al. (2006) performed linkage disequilibrium mapping of rachilla hair gene in barley and found that rachilla hair length was associated with SSR marker Bmag0223. Javaid and Taketa (2008) performed molecular mapping of srh using SSR and Expressed Sequence Tag (EST) markers on chromosome 5HL in barley by converting three monomorphic EST markers into polymorphic Cleaved Amplified Polymorphic Sequence (CAPS) markers. Short rachilla hair gene srh was mapped between two markers at a distance of 1.6 and 0.5 cm in the proximal and distal side, respectively.
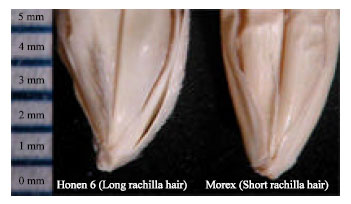 | |
| Fig. 1: | Long rachilla hair in Honen 6 (left) and short rachilla hair in Morex (right) |
Efforts are being carried out for fine mapping of short rachilla hair gene using different techniques such as association genetics (Waugh et al., 2009). The present research was conducted to refine the mapping of short rachilla hair gene srh in barley using 110 F2 plants and 320 recessive homozygous plants that were segregated in F2 generation of a different barley population. Moreover, using linked flanking markers k06288KU and Bmag0223, genetic diversity in world barley germplasm collection of short and long rachilla hair varieties was studied to understand the origin of short rachilla hair gene.
MATERIALS AND METHODS
A segregating F2 population of 110 plants was developed from the cross combination Morex/Honen 6 and was planted in the greenhouse. The F3 progeny test was conducted by planting twenty F3 plants from each F2 plant in the field during winter 2008-2009 to estimate the genotypes of F2 plants. Rachilla hair type was recorded in the F2 population from Morex/Honen 6 and the F3 population planted for progeny test. On the basis of progeny test, genotype of F2 plants was determined. A large population of 1318 F2 plants from Morex/Honen 6 was planted in the field and 320 homozygous recessive short rachilla hair (srh) plants were selected. Barley germplasm consisting of 90 short rachilla hair and 102 long rachilla hair accessions from different regions of the world was obtained from Research Institute for Bioresources, Okayama University, Japan and National Plant Germplasm System (NPGS), USA (Table 1). Germplasm was grown in the field for rachilla hair data recording and DNA extraction.
DNA was isolated from fresh leaf tissues of each plant from segregating populations and germplasm accessions using a modified form of SDS-based method (Csaikli et al., 1998) and analyzed for CAPS marker k06288KU (Javaid and Taketa, 2008) and SSR marker Bmag0223 (Ramsay et al., 2000).
| Table 1: | Origin of barley germplasm used |
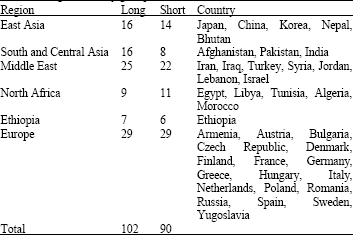 | |
For CAPS marker k06288KU analysis, DNA templates were amplified u sing EST primers k06288F 5’ ATACGCAACAAAGCGGACAC 3’ and k06288R 5’ TGGTTGGTCGGTCGTTTACT 3’. PCR of each DNA sample was conducted in a total volume of 15 μL containing 0.015 units AmpliTaq Gold DNA Polymerase (Applied Biosystems), 1X PCR buffer, 0.3 μM each primer, 0.2 mM dNTP, 1.3 mM MgCl2 and 10 ng template DNA. Thermal Cycler PTC-100 of MJ Research was used and program was set with an initial denaturing temperature of 94°C for 10 min followed by 30 cycles of denaturing temperature of 94°C for 1 min, annealing temperature of 54°C for 1 min and extension temperature of 72°C for 2 min. Final extension temperature was set at 72°C for 10 min. The PCR products were digested at 37°C for 4 h with restriction enzyme XspI. The digestion was conducted in 12 μL reaction mixture containing 10 μL PCR product and 1.6 U restriction enzyme in Takara 10xreaction buffer provided with the enzyme.
The SSR marker Bmag0223 was amplified using forward primer 5’ TTAGTCACCCTCAACGGT 3’ and reverse primer 5’ CCCCTAACTGCTGTGATG 3’, as reported by Ramsay et al. (2000). The PCR mixture for SSR analysis contained 0.01 units AmpliTaq Gold DNA Polymerase, 1X PCR buffer, 0.3 μM primers, 0.2 mM dNTP, 1.5 mM MgCl2 and 20 ng template DNA in a total volume of 10 μL. An initial 10 min denaturation at 94°C was followed by 30 cycles of 1 min denaturation at 94°C, 1 min annealing at 55°C and 1 min extension at 72°C. A final 5 min extension at 72°C completed the program. The PCR was performed in Thermal Cycler PTC-100 of MJ research.
The PCR products from mapping population of 110 F2 plants were resolved by agarose gel electrophoresis using 2.0% agarose gels containing ethidium bromide for Bmag0223 and 1.5% gels for k06288KU. Acrylamide gel electrophoresis using 10% acrylamide gels with SYBR safe staining was used to resolve the PCR products from 320 homozygous recessive short rachilla hair plants and germplasm collection of 192 plants.
Linkage analysis among srh locus, Bmag0223 and k06288KU was performed for each of 110 F2 plants and 320 homozygous recessive plants using the software Mapmaker 3.0 (Lander et al., 1987). The Kosambi mapping function was used to convert recombination frequencies into map distance (Kosambi, 1944). Combined data of 526 plants were analyzed including 96 F2 plants described previously and reported by Javaid and Taketa (2008).
Molecular analysis of 192 germplasm accessions was conducted using CAPS marker k06288KU and SSR marker Bmag0223. The k06288KU PCR products after digestion with XspI were classified into either Morex-type (without restriction sites, recorded as A) or Honen 6-type (with a restriction site, written as B). For Bmag0223, PCR product size showed continuous variation, thus, threshold size was determined at 145 bp (the size of short rachilla-type Morex); accessions with PCR product size with 145 bp or longer were considered as Morex-type or A, while, those with a shorter size were recorded as Honen 6-type or B. Allele frequency of flanking markers in short and long rachilla hair germplasm accessions was determined for different geographical regions through pie charts. Genetic diversity in each region was estimated by Shannon index (Lattoo et al., 2008).
RESULTS AND DISCUSSION
Mapping srh in F2 population: Genotyping of F2 population of 110 plants using k06288KU (Fig. 2) and Bmag0223 (Fig. 3) revealed the presence of six recombinants for k06288KU and two recombinants for Bmag0223. The srh locus was mapped between flanking markers k06288KU and Bmag0223 at 2.8 cm proximal and 0.8 cm distal side to the srh, respectively (Fig. 4a, b).
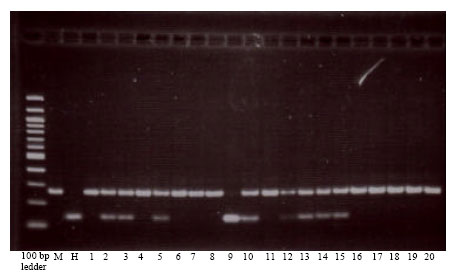 | |
| Fig. 2: | Genotyping of Morex (M) and Honen 6 (H) and F2 population of Morex/Honen 6 using CAPS marker k06288KU |
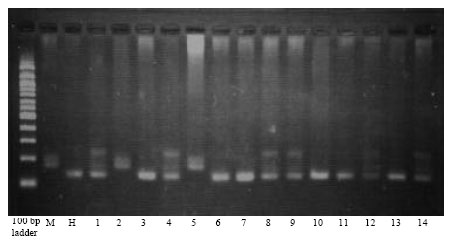 | |
| Fig. 3: | Genotyping of Morex (M), Honen 6 (H) and F2 population Morex/Honen 6 using SSR marker Bmag0223 |
Linkage analysis in homozygous recessive plants: Linkage analysis through genotyping of 320 homozygous recessive srh plants for k06288KU and Bmag0223 showed 39 recombinants for Bmag0223 while 5 recombinants were found for k06288KU. One plant showed recombination for both markers. Genetic map constructed through linkage analysis of srh with k06288KU and Bmag0223 mapped the markers at a genetic distance of 0.8 and 6.1 cm, respectively (Fig. 4c). An integrated map was constructed using the data from 206 F2 and 320 homozygous recessive plants in which k06288KU was mapped 1.6 cm proximal to srh while Bmag0223 had distance of 4.0 cm on distal side of srh (Fig. 4d).
Germplasm evaluation: Germplasm analysis using CAPS marker k06288KU divided the germplasm into 96 accessions with non restricted PCR products (allele A) including 32 short rachilla hair and 64 long rachilla hair accessions and 96 accessions with restricted PCR products (allele B) out of which 58 are short rachilla hair and 38 were long rachilla hair. The PCR analysis of germplasm using SSR marker Bmag0223 indicated that 130 germplasm accessions had long PCR products (allele A) similar to Morex including 72 short rachilla hair and 58 long rachilla hair accessions while 62 accessions had short PCR products (allele B) similar to Honen 6 with 18 short rachilla hair and 44 long rachilla hair accessions (Fig. 5).
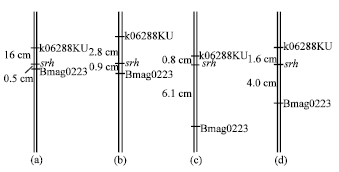 | |
| Fig. 4: | (a) Genetic map of short rachilla hair and molecular markers on chromosome 5HL constructed by Javaid and Taketa (2008) using 96 F2 plant from Morex/Honen 6. (b) Map of srh locus and flanking markers constructed using 110 F2 from Morex/Honen 6. (c) Map of srh locus and flanking markers constructed using 320 homozygous recessive srh plants from Morex/Honen 6. (d) Combined map of srh locus and flanking markers constructed using collective data of 526 plants including 206 F2 plants and 320 homozygous recessive short rachilla hair plants from Morex/Honen 6 |
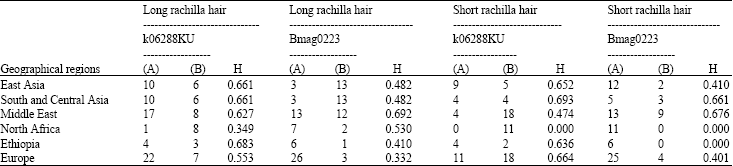
Diversity index was calculated in long and short rachilla hair germplasm accessions on the basis of frequency of different alleles of k06288KU and Bmag0223 (Table 2). In case of long rachilla hair accessions, diversity index for k06288KU ranged from 0.661 (East Asia and South and Central Asia) to 0.349 (North Africa), while, highest Bmag0223 diversity index was found in Middle East (0.692) and lowest in Europe (0.332). Short rachilla hair accessions showed highest diversity index for k06288KU in South and Central Asia and no diversity in North Africa. Bmag0223 diversity in short rachilla hair ranged from 0.676 (Middle East) to zero in North Africa and Ethiopia.
 | |
| Fig. 5: | Genotyping of Morex, Honen 6 and barley germplasm accessions with short and long rachilla hair from different geographical regions of the world using SSR marker Bmag0223 |
| Table 2: | Frequency and diversity index (H) of flanking markers k06288KU and Bmag0223 in long and short rachilla hair germplasm from different geographical regions |
 | |
A and B in parentheses show allele type of each marker, where A is Morex (short rachilla) type and B is Honen 6 (long rachilla) type | |
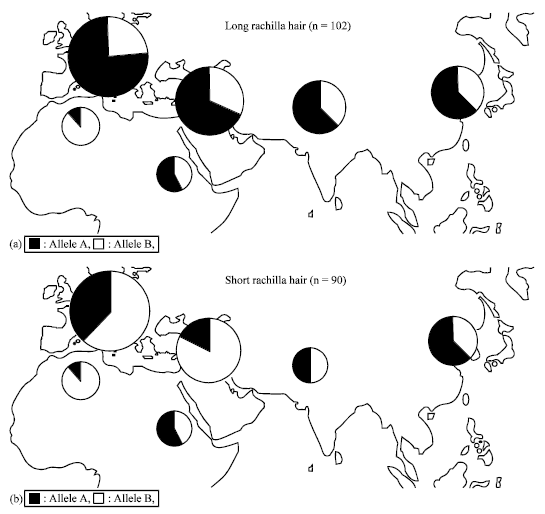
Results of germplasm analysis using k06288KU are summarized in Fig. 6a and b. Barley accessions were divided into short rachilla hair and long rachilla hair. No clear regional difference in allele frequency was found except for North Africa, where, the frequency of allele B is high both in long and short rachilla accessions. In other regions excluding Europe and Middle East, alleles A and B were almost equally represented. In Europe and Middle East, allele A was high in long rachilla group and allele B was high in short rachilla group.
Bmag0223 analysis showed regional difference in allele frequency. Although, allele A and B was almost equally represented in Middle East regardless of rachilla type, allele A became predominant in Europe, North Africa and Ethiopia regardless of rachilla type. On the other hand, in the Eastern regions, allele A tends to be associated with long rachilla phenotype and allele B is associated with short rachilla phenotype. This association h as become most evident in East Asia.
Short rachilla hair srh was mentioned in many genetic linkage maps constructed on the basis of morphological and molecular markers. The SSR marker Bmag0223 is one of the most important molecular markers for the study of short rachilla hair. It is found less than 3 cm distal to short rachilla hair gene in different mapping populations (Kraakman et al., 2006). Sato et al. (2004) constructed a large scale map of ESTs on barley genome and reported a genetic distance of 2.2 cm between k06288 and Bmag0223. Javaid and Taketa (2008) analyzed EST markers on chromosome 5HL and developed CAPS markers. In earlier study using 96 F2 plants, short rachilla hair locus srh was mapped between EST derived CAPS marker k06288KU and Bmag0223 within a genetic distance of 2.1 cm (Fig. 7a, b).
 | |
| Fig. 6: | Relationship between rachilla type and k06288KU polymorphism in 192 barley accessions from different regions of the world. (a) Upper map is for long rachilla accessions and (b) lower map is for short rachilla accessions |
The mapping in this experiment using F2 population of 110 plants shows similar results to previous reports. Bmag0223 is found to be the most closely linked molecular marker to short rachilla hair gene with a genetic distance of 0.9 cm distal to srh followed by k06288KU at a distance of 2 .8 cm proximal to srh. A population of 320 homozygous recessive srh plants gave results that are contrary to earlier reports. Integrated map constructed using data from 526 plants mapped k06288KU 1.6 cm proximal and Bmag0223 4.0 cm distal to srh. This difference of results may be, due to the population size. During the earlier experiments, smaller populations with few homozygous recessive srh plants were used, while, large population of 320 homozygous recessive srh plants was used during this experiment. This improvement in mapping technique has shown high recombination of Bmag0223 in homozygous recessive srh plants.
Barley is differentiated into Oriental and Occidental cultivars on the basis of morphological characters arisen by mutation under domestication and molecular markers. Short rachilla hair is one of the most important morphological characters that show a difference between barley germplasm from two geographical regions (Takahashi, 1955). Kahler and Allard (1981) examined worldwide pattern of genetic variation among isozyme loci and reported the classification of germplasm in the oriental and occidental groups. Strelchenko et al. (1999) confirmed the oriental-occidental differentiation of domesticated barley using RAPD markers. During this experiment, there is a clear demarcation in the germplasm for SSR marker Bmag0223. Allele A is high in West, while, allele B is high in East. High frequency of both alleles is found in Middle East. Thus, geographical differentiation of barley and high diversity in the Middle East is substantiated by SSR marker Bmag0223.
Study of the origin of morphological characters on the basis of molecular analysis, sometimes, leads to multiple origin such as origin of six-rowed phenotype through independent mutations of vrs1 (Komatsuda et al., 2007) while, for some characters monophyletic origin is reported like origin of naked caryopsis gene nud (Taketa et al., 2008). Evaluation of long rachilla hair germplasm using Bmag0223 shows a clear geographical differentiation, while, no useful information was obtained from a more closely-linked marker k06288KU. Results of Bmag0223 show a different transition of alleles from West to East.
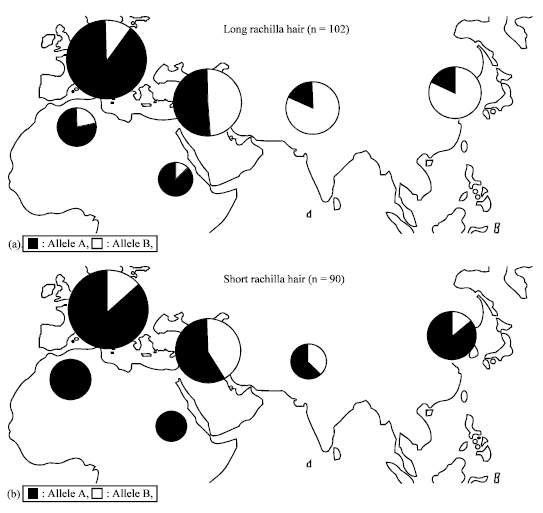 | |
| Fig. 7: | Relationship between rachilla type and Bmag0223 polymorphism in 192 barley accessions from different regions of the world. (a) Upper map is for long rachilla accessions and (b) lower map is for short rachilla accessions |
Frequencies of both alleles were similar in Middle East. North Africa, Ethiopia and Europe are represented by high frequencies of Bmag0223 allele A regardless of rachilla types, while, in East Asia and South and Central Asia allele A is associated with short rachilla and allele B is associated with long rachilla. The transition did not occur in Middle East, which is considered a center of origin in barley (Badr et al., 2000) but, it became evident as barley spread to west and east. We suppose that somewhere in Middle East, short rachilla mutation occurred in accessions with Bmag0223 allele A and allele A type of both long and short rachilla type predominantly spread westward. On the contrary, to the eastward direction, long rachilla accessions with allele B and short rachilla accessions with allele A appear to have moved. Regional differences observed in Bmag0223 alleles closely linked to the srh locus on 5HL seem to support occidental and oriental differentiation in domesticated barley. Further studies using more closely linked molecular markers near the srh locus may give clearer picture about the origin and migration route of domesticated barley.
ACKNOWLEDGMENT
The authors are grateful to Dr. K. Sato, Okayama Univeristy, Japan for providing primers for EST-based markers on 5HL and barley germplasms.
REFERENCES
- Badr, A., K. Mueller, R. Schaefer-Pregl, H. El-Rabey and S. Efgen et al., 2000. On the origin and domestication history of barley (Hordeum vulgare). Mol. Biol. Evol., 17: 499-510.
PubMedDirect Link - Costa, J.M., A. Corey, M. Hayes, C. Jobet and A. Kleinhofs et al., 2001. Molecular mapping of the Oregon Wolfe Barleys: A phenotypically polymorphic doubled-haploid population. TAG Theor. Applied Genet., 103: 415-424.
CrossRefDirect Link - Csaikl, U.M., H. Bastian, R. Brettschneider, S. Gauch and A. Meir et al., 1998. Comparative analysis of different DNA extraction protocols: A fast, universal maxi-preparation of high quality plant DNA for genetic evaluation and phylogenetic studies. Plant Mol. Biol. Rep., 16: 69-86.
CrossRefDirect Link - Franckowiak, J.D., 1997. Revised linkage maps for morphological markers in barley, Hordeum vulgare. Barley Genet. News, 26: 9-21.
Direct Link - Kahler, A.L. and R.W. Allard, 1981. Worldwide pattern of genetic variation among four esterase loci in barley (Hordeum vulgare L.). Theor. Applied Genet., 59: 101-110.
CrossRefDirect Link - Kikuchi, S., S. Taketa, M. Ichii and S. Kawasaki, 2003. Efficient fine mapping of the naked caryopsis gene (nud) by HEGS (High efficiency genome scanning) AFLP in barley. Theor. Applied Genet., 108: 73-78.
CrossRefDirect Link - Kebebew, F., Y. Tsehaye and T. McNeilly, 2001. Morphological and farmers cognitive diversity of barley (Hordeum vulgare L. [Poaceae]) at Bale and North Shewa of Ethiopia. Genet. Res. Crop Evol., 48: 467-481.
CrossRefDirect Link - Kleinhofs, A., A. Kilian, M.A.S. Maroof, R.M. Biyashev and P. Hayes et al., 1993. A molecular, isozyme and morphological map of the barley (Hordeum vulgare) genome. Theor. Applied Genet., 86: 705-712.
CrossRefDirect Link - Komatsuda, T. and Y. Mano, 2002. Molecular mapping of the intermedium spike-c (int-c) and non-brittle rachis 1 (btr1) loci in barley (Hordeum vulgare L.). Theor. Applied Genet., 105: 85-90.
CrossRefDirect Link - Komatsuda T., M. Pourkheirandish, C. He, P. Azhaguvel and H. Kanamori et al., 2007. Six-rowed barley originated from a mutation in a homeodomain-leucine zipper I-class homeobox gene. Proc. Nat. Acad. Sci., 104: 1424-1429.
CrossRefPubMedDirect Link - Komatsuda, T., P. Maxim, N. Senthil and Y. Mano, 2004. High-density AFLP map of nonbrittle rachis 1 (btr1) and 2 (btr2) genes in barley (Hordeum vulgare L.). Theor. Applied Genet., 109: 986-995.
CrossRefPubMedDirect Link - Kraakman, A.T.W., F. Martinez, B. Mussiralie, F.A.V. Eeuwijk and R.E. Niks, 2006. Linkage disequilibrium mapping of morphological, resistance and other agronomically relevant traits in modern spring barley cultivars. Mol. Breed., 17: 41-58.
CrossRefDirect Link - Lander, E.S., P. Green, J. Abrahamson, A. Barlow, M.J. Daly, S.E. Lincoln and L. Newburg, 1987. MAPMAKER: An interactive computer package for constructing primary genetic linkage maps of experimental and natural populations. Genomics, 1: 174-181.
CrossRefDirect Link - Lattoo, S.K., R.S. Dhar, S. Khan, S. Bamotra, M.K. Bhan, A.K. Dhar and K.K. Gupta, 2008. Comparative analysis of genetic diversity using molecular and morphometric markers in Andrographis paniculata (Burm. f.) Nees. Genet. Res. Crop Evol., 55: 33-43.
CrossRefDirect Link - Parzies, H.K., W. Spoor and R.A. Ennos, 2000. Genetic diversity of barley landrace accessions (Hordeum vulgare sp. vulgare) conserved for different lengths of time in ex situ gene banks. Heredity, 84: 476-486.
PubMed - Ramsay, L., M. Macaulay, S.D. Ivanissevich, K. MacLean and L. Cardle et al., 2000. A simple sequence repeat-based linkage map of barley. Genetics, 156: 1997-2005.
PubMedDirect Link - Salamini, F., H. Ozkan, A. Brandolini, R. Schafer-Preg and W. Martin, 2002. Genetics and geography of wild cereal domestication in the near East. Nat. Rev. Genet., 3: 429-441.
CrossRef - Strelchenko, P., O. Kovalyova and O. Kazutoshi, 1999. Genetic differentiation and geographical distribution of barley germplasm based on RAPD markers. Genet. Resourc. Crop Evol., 46: 193-205.
CrossRefDirect Link - Taketa, S., S. Kikuchi, T. Awayama, S. Yamamoto, M. Ichii and S. Kawasaki, 2004. Monophyletic origin of naked barley inferred from molecular analyses of a marker closely linked to the naked caryopsis gene (nud). Theor. Applied Genet., 108: 1236-1242.
CrossRefDirect Link - Taketa, S., S. Amano, Y. Tsujino, T. Sato and D. Saisho et al., 2008. Barley grain with adhering hulls is controlled by an ERF family transcription factor gene regulating a lipid biosynthesis pathway. Proc. Nat. Acad. Sci., 105: 4062-4067.
CrossRefPubMedDirect Link - Tanno, K., S. Taketa, K. Takeda and T. Komatsuda, 2002. A DNA marker closely linked to the vrs1 locus (row-type gene) indicates multiple origins of six-rowed cultivated barley (Hordeum vulgare L.). Theor. Applied Genet., 104: 54-60.
CrossRefPubMedDirect Link - Sato, K., N. Nankaku, Y. Motoi and K. Takeda, 2004. A large scale mapping of ESTs on barley genome. Proceedings of the 9th International Barley Genetics Symposium, Vol. 1, Jun. 20-26, Brno, Czech Republic, pp: 79-85.
Direct Link