
A. Arunyanark
Department of Plant Science and Agricultural Resources, Faculty of Agriculture, Khon Kaen University, Muang, Khon Kaen 40002, Thailand
S. Jogloy
Department of Plant Science and Agricultural Resources, Faculty of Agriculture, Khon Kaen University, Muang, Khon Kaen 40002, Thailand
N. Vorasoot
Department of Plant Science and Agricultural Resources, Faculty of Agriculture, Khon Kaen University, Muang, Khon Kaen 40002, Thailand
C. Akkasaeng
Department of Plant Science and Agricultural Resources, Faculty of Agriculture, Khon Kaen University, Muang, Khon Kaen 40002, Thailand
T. Kesmala
Department of Plant Science and Agricultural Resources, Faculty of Agriculture, Khon Kaen University, Muang, Khon Kaen 40002, Thailand
A. Patanothai
Department of Plant Science and Agricultural Resources, Faculty of Agriculture, Khon Kaen University, Muang, Khon Kaen 40002, Thailand
Asian Journal of Plant Sciences
Year: 2009 | Volume: 8 | Issue: 2 | Page No.: 102-110







ABSTRACT
The objectives of this study were to examine the stability of SPAD (soil plant analysis development) chlorophyll meter reading (SCMR) and chlorophyll density, surrogate trait of drought tolerance in peanut (Arachis hypogaea L.) and their relationships in different leaf positions at different times under different drought stress conditions. Chlorophyll density and SCMR varied depending on water regimes, times of sampling and genotypes, but water regime x genotype interactions were not significant for chlorophyll density and SCMR. The correlation coefficients between chlorophyll density and SCMR were positive and significant across irrigation treatments (r = 0.76**, 0.94** and 0.96**) and each water regime, plant age and leaf position (r = 0.31 to 0.99**). Interestingly, chlorophyll density and SCMR at different water regimes were also correlated significantly, indicating the stability of their relationship in different water regime conditions. The result suggest that evaluation of chlorophyll density by SCMR can be carried out at any water regime conditions in the second or third-fully expanded leaves after 40 days of crop growth. This confers a large flexibility to application of SCMR in breeding program for drought tolerance.
PDF Abstract XML References Citation
How to cite this article
A. Arunyanark, S. Jogloy, N. Vorasoot, C. Akkasaeng, T. Kesmala and A. Patanothai, 2009. Stability of Relationship Between Chlorophyll Density and Soil Plant Analysis Development Chlorophyll Meter Readings in Peanut Across Different Drought Stress Conditions. Asian Journal of Plant Sciences, 8: 102-110.
DOI: 10.3923/ajps.2009.102.110
URL: https://scialert.net/abstract/?doi=ajps.2009.102.110
DOI: 10.3923/ajps.2009.102.110
URL: https://scialert.net/abstract/?doi=ajps.2009.102.110
INTRODUCTION
Crop productivity per unit of water has become an important consideration in breeding programs aimed at developing water use efficient (WUE) varieties. WUE or transpiration efficiency (TE, defined as the biomass produced per unit of water transpired) has been widely perceived as a useful trait for drought tolerance (Richards, 1996). However, it is difficult to measure TE, a complex physiological trait, in segregating populations and breeding lines in large-scale breeding programs. Further research has shown that TE is strongly correlated with the easily measurable trait i.e., specific leaf area (SLA) (leaf thickness) (Wright et al., 1993; Nageswara Rao and Wright, 1994; Wright and Nageswara Rao, 1994). Significant relationships between TE, SLA, chlorophyll and the photosynthetic enzyme ribulose 1-5 bisphosphate carboxylase (Rubisco) content in peanut suggested that photosynthetic capacity is the main cause of variation for TE in peanut (Nageswara Rao et al., 1995).
Chlorophyll content per unit leaf area (chlorophyll density) has been used as an index of photosynthetic capacity and growth of many crop plants and it is most important to crop performance because chlorophyll is a major photosynthetic pigment. The major role of this pigment is to absorb and reacts with visible light in the photosynthesis (Bowyer and Leegood, 1997). The ability to maintain high chlorophyll density under water deficit conditions has been suggested as a drought tolerance mechanism in barley (This et al., 2000) and potato (Van der Mescht et al., 1999). In peanut, Arunyanark et al. (2008) demonstrated that the variation in TE was closely correlated with genotypic variation in chlorophyll density and hence with photosynthetic capacity, such that chlorophyll density could be used as a potential indicator of TE in peanut.
Several methods with well-equipped laboratory have been available for assessing chlorophyll density. Chlorophyll is not soluble in water and it can be extracted in 80 % (v/v) acetone solution (Arnon, 1949) or N,N-dimethylformamide (DMF) (Moran, 1981) and then its content is determined by spectrophotometer. Although chlorophyll density is a drought tolerance trait, this measurement process is laborious, time consuming, costly, destructive and inconvenient for a large number of samples. These limitations preclude its use for plant breeding programs in which large numbers of samples are involved. Indirect methods which are easy and rapid to use, economical, effective and reliable to assess chlorophyll density are required.
Minolta SPAD-502 meter has been developed to assess crop health and becomes a useful tool for scientists conducting experiments in plant science. The SPAD meter measures green color intensity in leaves in vivo and is, therefore, an ideal instrument for obtaining the data without destructive sampling of a large number of chlorophyll data in the field and the reading is performed in very short time. Close associations between SPAD chlorophyll meter reading (SCMR) value and chlorophyll density had been reported in leaves of maize, soybean (Markwell et al., 1995), cotton (Wu et al., 1998), rice (Ommen et al., 1999), wheat and potato (Bindi et al., 2002; Uddling et al., 2007). In peanut, the information on the relationships between SCMR and chlorophyll density in leaves also was reported by Samdur et al. (2000) and Arunyanark et al. (2008). Although a close correlation between chlorophyll density and SCMR has been established, it can therefore be inferred that chlorophyll density-SCMR relationship might be influenced by factors such as time of observation, leaf age and water deficit conditions.
The use of SCMR to assess relative chlorophyll density as alternative to the standard method is very attractive because it is easy to operate, low cost, non-destructive technique and can be applied in the field conventionally. Moreover, SCMR is easy to measure, highly correlated with SLA and TE (Nageswara Rao et al., 2001; Sheshshayee et al., 2006; Arunyanark et al., 2008). Thus, SCMR could be used indirectly for evaluation of drought tolerance in peanut genotypes. However, breeding and selection scheme for drought tolerance was limited by other important problems. Such breeding programs have to be conducted under dry season only or in glasshouse where drought conditions can be simulated. In field evaluation, although the number of genotypes evaluated is not limited, it can be grown only one crop a year. In contrast to field evaluation, although the experiment in greenhouse can simulate drought, it has a limited space to grow a large number of peanut genotypes. Moreover, in large-scale breeding programs, it is difficult to complete SCMR observations within a specified time and plant stage, or could not take sample at same leaf position for all.
Surrogate traits for drought tolerance such as chlorophyll density and SCMR will help to increase potential of breeding scheme if they are able to identify drought tolerance genotypes with chlorophyll density and SCMR stability across water supply conditions. Thus, the evaluation can be carried out for all seasons. Moreover, SCMR would be helpful to breeders if these measurements could be observed in several times or several leaf positions. Therefore, information on the effects of different water regimes, plant ages and leaf positions on chlorophyll density and SCMR relationships is required.
The experiments with a wide range of peanut germplasm and environment conditions will help clarify the technique. The objectives of this study were to examine the stability of chlorophyll density and SCMR and evaluate the relationships between chlorophyll density and SCMR in different leaf positions at different times under different drought stress conditions. These observations can be applied for developing peanut varieties tolerant to drought stress conditions.
MATERIALS AND METHODS
Experimental design and treatments: The field experiment was conducted at the Field Crop Research Station, Department of Plant Science and Agricultural Resources, Faculty of Agriculture, Khon Kaen University (latitude 16° 26 N, longitude 102° 50 E), on Yasothon soil series (loamy sand, Oxic Paleustults), in Thailand, during the dry seasons (November 2003-May 2004). A split-plot in a randomized complete block design with four replications was used. Three different water regimes (field capacity; FC, 2/3 available water (AW) and 1/3 AW) were assigned in main plots and twelve peanut cultivars in sub-plots. Twelve peanut cultivars were chosen based on their differences in drought resistance and leaf color. Eight of these lines (ICGV 98300, ICGV 98303, ICGV 98305, ICGV 98308, ICGV 98324, ICGV 98330, ICGV 98348 and ICGV 98353) were drought resistant germplasm introduced from the International Crops Research Institute for Semi-Arid Tropics (ICRISAT). Tifton-8 is a drought resistant line introduced from the United State Department of Agriculture (USDA). Two cultivars (KK 60-3 and Tainan 9) were commercial varieties grown in Thailand and are known to be sensitive to drought. A non-nodulating line (yellow- green leaf color) was used as a control for low chlorophyll and drought susceptible genotype. The crop in each sub-plot (peanut cultivar) was grown in eleven-row plots, 6 m long, with a spacing of 0.5 m between rows and 0.2 m between plants within row. A subsoil-drip-irrigation system installed at 10 cm below the soil surface and fitted with a pressure valve and water meter ensured the uniform supply of measured amount of water across each plot. The more detailed experiment setup, irrigation control and crop management had been reported previously by Arunyanark et al. (2008).
Data collections
Soil moisture content: Soil moisture contents were measured by the gravimetric method at the depths of 0-5, 25-30 and 55-60 cm at planting and harvest. Weekly changes of soil moisture were measured by the neutron method from all sub-plots at 30, 60 and 90 cm of soil depth.
Chlorophyll density and SPAD chlorophyll meter readings: Data were recorded for SCMR, chlorophyll density (chlorophyll content per unit leaf area) at three times as 20, 40 and 60 (DAE). Five plants from each sub-plot were randomly sampled and the first, second and third fully-expanded leaves from the top of the main stems were used for SCMR assessment at 9.00-10.00 am, SCMR were recorded by a Minolta SPAD-502 meter (Tokyo, Japan) on the four leaflets of each individual leaf as described (Nageswara Rao et al., 2001). An average SCMR for each plot was derived from 20 single observations (4 leafletsx5 plants/plot).
Only one leaflet from each sample leaf in each plot was used for chlorophyll determination. The leaflet previously used for SCMR was harvested and kept chilled unit it was analyzed. One leaf disc was taken from each leaf let using a cork borer with a diameter of 1 cm2. Leaf discs were soaked in 5 mL of N,N-diamethylformamide and kept in dark for 24 h before determining chlorophyll using a light absorption with a spectrophotometer.
Chlorophyll density (per unit leaf area) was analyzed following the procedures described by Moran (1981).
Statistical analysis: The data were pooled over times of sampling to investigate the main effects and interaction effects. The pooled analysis of variance for measurements over time based on a split plot design (Gomez and Gomez, 1984) was performed for SCMR and chlorophyll density. Leaf positions were analyzed separately.
As time of sampling is the large source of interactions, separate times of sampling were later analyzed using a split plot design. Water regimes were assigned in main plots and peanut genotypes in subplots. Leaf positions were treated as sub-samples. Mean separations were conducted by using least significant difference (LSD). All calculations were conducted by PROC GLM in SAS (SAS Institute Inc., Cary, NC, USA).
Simple correlations among and within main effects under study were computed for SCMR and chlorophyll density.
RESULTS AND DISCUSSION
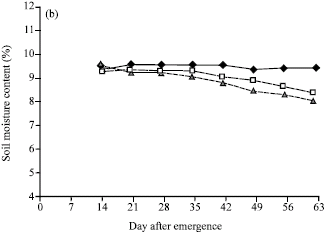
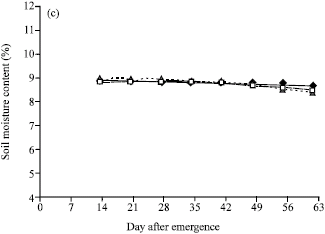
Soil moisture: Soil moisture levels for different water regimes, as determined by percentages of available moisture content, at different depths of the soil profile showed a reasonable control of the water-regime treatments. Clear differences in soil moisture percentages among water-regimes were shown, with soil moisture levels for the individual treatments being close to the expected percentages (10.55 , 8.48 and 6.40 % for FC, 2/3 AW and 1/3 AW) as determined by pressure plate from the soil properties. The differences among water regimes in soil moisture contents were much clearer at the depth of 30 cm in which soil moisture of 1/3 AW were lower than those of FC and 2/3 AW and soil moistures of 2/3 AW were lower than those of FC (Fig. 1a). Soil moisture at 60 cm of soil depth has clear difference between FC and water stress levels, but soil moisture contents at 2/3 AW and 1/3 AW levels were rather close together. At the depth of 90 cm, the soil moisture contents among three water regimes were overlapped (Fig. 1b, c).
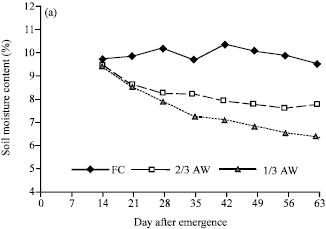 | |
| Fig. 1: | Soil moisture content (%) in three available water regimes (field capacity (FC), 2/3 available water (2/3 AW) and 1/3 available water (1/3 AW)) at (a) 30, (b) 60 and (c) 90 cm of the soil depth in the field |
G x E interaction effects for chlorophyll density and SCMR: Across times of sampling, differences in SCMR were found among water regimes in the first and second leaf positions, but the differences in chlorophyll density were not significant for all leaf positions (Table 1). The differences were also found among peanut genotypes and times of sampling in all leaf positions. The interaction effects associated with time of sampling (W x T and G x T) were most significant, whereas other interaction effects (G x W and G x W x T) were not significant for most cases except for G x W x T for chlorophyll density in second leaf position. Generally, G x E interactions was more pronounced for chlorophyll density than for SCMR, suggesting the stability of SCMR trait across environments, compared to chlorophyll density.
It is interesting to note here that, although water regimes and peanut genotypes were significantly different, their interaction effects were not significant for both chlorophyll density and SCMR (Table 1). This pattern was found in all leaf positions. The findings indicate the stability of SCMR and chlorophyll density in peanut across water regimes and peanut genotypes.
Similar to present findings, chlorophyll density and SCMR has shown to be a stable parameter across seasons and a low genotypexseason interaction was reported by Arunyanark et al. (2008) and Nageswara Rao et al. (2001). The results also suggested that selection for chlorophyll density and SCMR could be carried out under any water conditions under investigation with similar results. Timexgenotype interaction variances for chlorophyll density and SCMR, though significant, were much smaller than those of their associated main effects. The variations in chlorophyll density and SCMR across times of sampling were also affected by the differences in growth stages in which plant development was not complete at early growth stages. The higher G x E interaction variances in first leaf position than in second and third leaf positions also supports these findings and also suggests to avoid sampling at first leaf position.
Effects of drought stress on chlorophyll density and SCMR: Both chlorophyll density and SCMR gave similar information on the effects of drought stress in peanut. Differences in chlorophyll density and SCMR among water regimes were found at 60 DAE only and the highest chlorophyll density and SCMR were found at 1/3 AW (Table 2). The result indicates that more stress higher is the density of chlorophyll. Drought stress increased chlorophyll density as well as increasing transpiration efficiency in peanut (Arunyanark et al., 2008). The increase in chlorophyll density is most likely due to the thicker leaves of stressed plants (lower specific leaf area) (Nageswara Rao and Wright, 1994), causing higher chlorophyll content per unit area.
The peanut genotypes showed significant differences in chlorophyll density and SCMR with similar rankings at all sampling times (Table 2). ICGV 98324, ICGV 98330 and KK 60-3 gave high chlorophyll density and SCMR, Tifton-8, ICGV 98305, ICGV 98348 and ICGV 98353 showed intermediate, whereas non-nod was the lowest at all sampling times. The results suggested that ICGV 98324, ICGV 98330 and KK 60-3 are promising genotypes for use as parents in drought tolerance in breeding programs. There was an evidence that leaf chlorophyll density was used as a criterion in selection for TE in peanut (Arunyanark et al., 2008; Sheshshayee et al., 2006) and wheat (Fotovat et al., 2007). Therefore, ability to maintain chlorophyll density of water stressed plants can be a criterion in selection for drought tolerance. This et al. (2000) found that the measurement of the chlorophyll density could be considered as a simple quantitative trait related to drought tolerance in barley. Moreover, Van der Mescht et al. (1999) found that the levels of chlorophyll density could be used as an indication of drought tolerance in potato.
Differences in chlorophyll density and SCMR among leaf positions were found as early as 20 DAE and were likely to rank in the order of first < second < third, respectively. Peanut genotypes were also different in stages of development in which the maximum chlorophyll density in leaves was developed. Maximum chlorophyll density is dependent on plant ages and leaf ages and both characters are under genetic and physiological controls. This evidence indicated that Minolta SPAD-502 meter was able to detect difference in relative chlorophyll density even among leaves within peanut cultivars and therefore, sensitive in vivo measurement of relative chlorophyll density in peanut leaves.
Relationship between chlorophyll density and SCMR: The correlation coefficients between chlorophyll density and SCMR were positive and significant across irrigation treatments, with the correlation coefficients of 0.76**, 0.94** and 0.96** at 20, 40 and 60 DAE, respectively (Fig. 2). The results indicated a close and stable relationship between chlorophyll density and SCMR across water regimes in all plant ages.
Correlation coefficients between SCMR and chlorophyll density in peanut leaves were calculated for each water regime, plant age and leaf position (Table 3). Under well-watered conditions at 20 DAE, the correlation coefficients of three leaf positions ranging from r = 0.31 to 0.75** were much lower than those at 40 (r = 0.77** to 0.99**) and 60 (r = 0.93** to 0.97**) DAE.
| Table 1: | Pooled analysis of variance for measurements over time in split-plot design of chlorophyll density and SPAD chlorophyll meter reading (SCMR) |
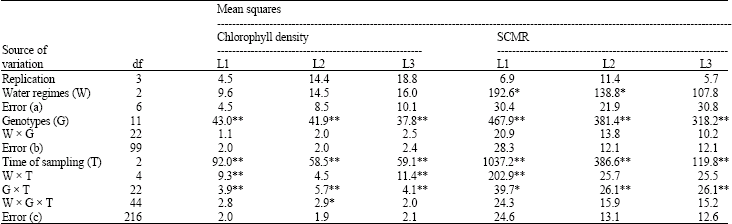 | |
| Significant at *p < 0.05 and **p < 0.01 levels. L1, L2 and L3: First, second and third leaf positions | |
| Table 2: | Chlorophyll density and SPAD chlorophyll meter reading (SCMR) of peanut at 20, 40 and 60 day after emergence (DAE) |
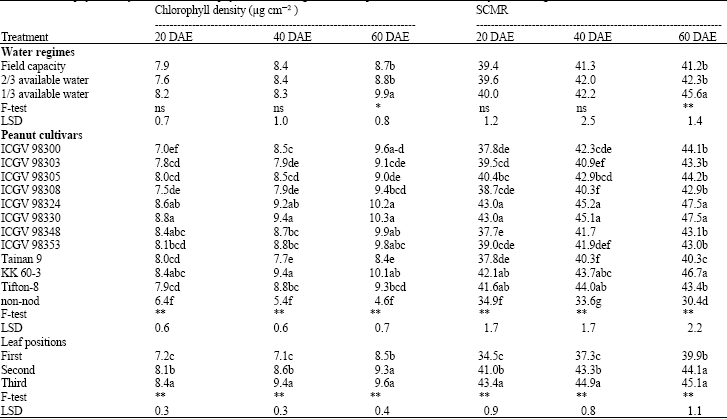 | |
| Mean in the same column within the same letters(s) with the same letters are not significantly different by least significant difference (LSD) at p < 0.05 | |
| Table 3: | Correlation coefficients between chlorophyll density and SPAD chlorophyll meter reading |
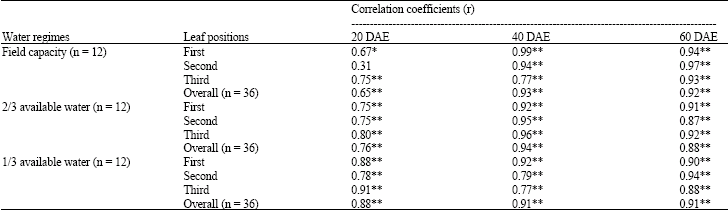 | |
| *, **Significant at the 0.05 and 0.01 probability level. DAE: Day after emergence | |
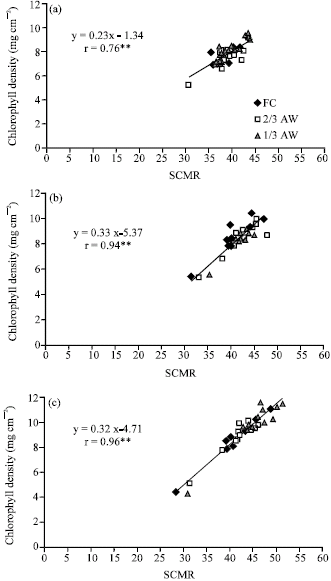 | |
| Fig. 2: | Correlation between chlorophyll density and SPAD chlorophyll meter reading (SCMR) across three water regimes (field capacity (FC), 2/3 available water (2/3 AW) and 1/3 available water (1/3 AW)) (n = 36) at (a) 20, (b) 40 and (c) 60 days after emergence. ** Significant at 0.01 probability level |
Under 2/3 AW, the correlation coefficients (r = 0.75** to 0.80**) at 20 DAE were significant and lower than those at 40 (r = 0.92** to 0.96**) and 60 (r = 0.87** to 0.92**) DAE. Under 1/3 AW, the correlation coefficients (r = 0.78** to 0.91**) at 20 DAE were generally lower than those at 40 (r = 0.77** to 0.92**) and 60 (r = 0.88** to 0.94**) DAE except for the third leaf position at 40 DAE (r = 0.77**).
The first-fully expanded leaf from the top had high correlation between chlorophyll density and SCMR. However, the photosynthetic machinery in the leaves in this position may not fully develop as indicated by lower chlorophyll density. Chlorophyll density in the first fully expanded leaf was always lower than that in the second and third fully expanded leaves. The variations in ontogenetic development of the first-fully expanded leaves may occur in peanut population. Therefore, precaution should be taken for determining chlorophyll density in peanut leaves. Present results indicated that the second or third fully expanded leaf would be more suitable for taking chlorophyll measurements or SPAD readings. Although there was no detail on justification for leaf samplings, Nautiyal et al. (2002) used leaflets of the second leaf from the apex of main stem of peanut cultivars for measurements of carbon exchange, relative water content and chlorophyll fluorescence.
When only sampling times at different water levels were considered, ignoring the effect of leaf position, the correlation coefficients between SCMR and chlorophyll density were highly significant (p < 0.01) at all of three water regimes (Table 3). However, at 20 DAE, the correlation coefficients (r = 0.65** to 0.88**) were lower than those at 40 (r = 0.91** to 0.94**) and 60 (r = 0.88** to 0.92**) DAE.
The correlation coefficients between SCMR and chlorophyll density were positive and highly significant, indicating closer relationship of SCMR and chlorophyll density in the first, second and third-fully expanded leaves and all the plant ages of sampling times, both at non-water stress and water stress conditions, except at 20 DAE when the lowest relationships were found.
The recent study suggested that water stress and different leaf positions had no significant effect on the relationship between SCMR and chlorophyll density and plant ages had a small effect on this relationship. The older plants the stronger correlation coefficients between SCMR and chlorophyll density would be. The results indicated that 40 and 60 DAE were the most suitable dates for taking chlorophyll measurement by SCMR because at these dates the correlation coefficient values were the highest. Moreover, at 40 and 60 DAE, maximum chlorophyll in leaves was developed as indicated by the highest chlorophyll density in these date. Sampling at 20 DAE would be less suitable due to its low correlation between chlorophyll density and SCMR. Moreover, the lowest chlorophyll density at 20 DAE indicated that chlorophyll was not developed completely at this age.
The SCMR is an indicator of the light-transmittance characteristics of the leaf, which is dependent on the leaf chlorophyll density (Richardson et al., 2002). The significant and positive correlation between SCMR and chlorophyll density across genotypes under a range of drought stress conditions in the present study (Table 3) demonstrated that SCMR could be a rapid tool to assess genotypic variation for leaf chlorophyll density in order to identify drought tolerant genotypes. Relationships between chlorophyll density and SCMR in wheat flag leaves (Ommen et al., 1999) and in potato leaves (Bindi et al., 2002) were curvilinear. In this study, there was a linear relationship between chlorophyll density and SCMR in peanut (Fig. 2 and 3c).
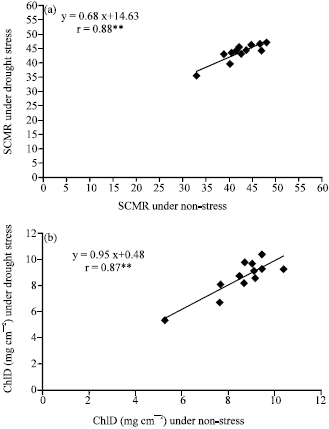 | |
| Fig. 3: | Correlation between chlorophyll density (ChlD) under non-stress (FC) and water stress (2/3 AW) conditions (a), between SPAD chlorophyll meter reading (SCMR) under non-stress and water stress conditions (b) and between SCMR under non-stress and ChlD under water stress conditions (c) in the second leaf position at 40 day after emergence. **Significant at 0.01 probability levels |
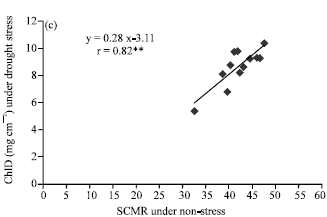
Interestingly, correlation between chlorophyll density under non-stress (FC) and water stress (2/3 AW) conditions (Fig. 3), between SCMR under non-stress and water stress conditions and between SCMR under non- stress and chlorophyll density under water stress conditions were strong, positive and significant in the second leaf position at 40 DAE. As well as, high significant correlation with each other were found for all combinations of water regimes both 40 and 60 DAE and with third leaf position (data not shown). It implied that chlorophyll density and SCMR, a surrogate trait of drought tolerance in peanut, were stable in different water regime conditions. It suggested that performance of chlorophyll density and SCMR were repeatable under both drought and non-drought stress conditions. This gives a large flexibility to breeders who want to evaluate genotypes for drought tolerance. Because the evaluation can be carried out without conducting drought conditions, it can be done for several crops a year.
However, light intensity has been shown to affect the chlorophyll meter readings by the Minolta SPAD-502 chlorophyll meter. Irradiance during the sampling period could also be critical. Hoel and Solhaug (1998) found that the SPAD reading was lowest in the middle of the day and highest at dusk and dawn. In addition, several factors affecting SPAD values consist of radiation between seasons, plant density, varietal groups, nutrient status in soil and biotic and abiotic stresses that induce leaf discoloration (Turner and Jund, 1994). Users should be aware of these interfering factors and should take adequate precaution against them while using the SCMR.
It has been concluded that plant age, drought stress and leaf positions could affect chlorophyll density and SCMR. However, water regime x genotype interaction was not significant for both chlorophyll density and SCMR, indicating the stability of both chlorophyll density and SCMR across different water regimes. The relationships between chlorophyll density and SCMR were high, positive and significant, suggesting that SCMR is a useful tool for assessing chlorophyll density in peanut. Plant ages had a small effect on the relationships between chlorophyll density and SCMR, whereas water stress and leaf positions had no significant effect on the relationship. It is clear that using SCMR for evaluation of chlorophyll density can be recorded at any water regime conditions in the second or third-fully expanded leaves after 40 days of crop growth. This gives a large flexibility to breeders who have to record observations in a large number of segregating and breeding populations in the field. Drought simulation is not necessary because selection under drought condition gives the results similar to those under well watered condition.
ACKNOWLEDGMENTS
The authors are grateful for the financial support of the Royal Golden Jubilee Ph.D Program (Grant No. PHD/0026/2547) and the Senior Research Scholar Project of Dr. Aran Patanothai under the Thailand Research Fund and also supported in part by the basic research for supporting groundnut varietals improvement for drought tolerance project under the Thailand Research Fund.
REFERENCES
- Arnon, D.I., 1949. Copper enzymes in isolated chloroplasts. Polyphenoloxidase in Beta vulgaris. Plant Physiol., 24: 1-15.
CrossRefPubMedDirect Link - Arunyanark, A., S. Jogloy, C. Akkasaeng, N. Vorasoot and T. Kesmala et al., 2008. Chlorophyll stability is an indicator of drought tolerance in peanut. J. Agron. Crop Sci., 194: 113-125.
CrossRefDirect Link - Bindi, M., A. Hacour, K. Vandermeiren, J. Craigon, K. Ojanpera, G. Sellden, P. Hogy, J. Finnan and L. Fibbi, 2002. Chlorophyll concentration of potatoes grown under elevated carbon dioxide and/or ozone concentrations. Eur. J. Agron., 17: 319-335.
CrossRefDirect Link - Fotovat, R., M. Valizadeh and M. Toorchi, 2007. Association between water-use-efficiency components and total chlorophyll content (SPAD) in wheat (Triticum aestivum L.) under well-watered and drought stress conditions. J. Food. Agric. Environ., 5: 225-227.
Direct Link - Gomez, K.A. and A.A. Gomez, 1984. Statistical Procedures for Agricultural Research. 2nd Edn., John Wiley and Sons Inc., Hoboken, New Jersey, ISBN: 978-0-471-87092-0, Pages: 704.
Direct Link - Hoel, B.O. and K.A. Solhang, 1998. Effect of irradiance on chlorophyll estimation with the Minolta SPAD-502 leaf chlorophyll meter. Ann. Botany, 82: 389-392.
Direct Link - Markwell, J., J.C. Osterman and J.L. Mitchell, 1995. Calibration of the Minolta SPAD-502 leaf chlorophyll meter. Photosynth. Res., 46: 467-472.
Direct Link - Moran, R., 1982. Formulae for determination of chlorophyllous pigments extracted with N,N-dimethylformamide. Plant Physiol., 69: 1376-1381.
CrossRefDirect Link - Nageswara Rao, R.C. and G.C. Wright, 1994. Stability of the relationship between specific leaf area and carbon isotope discrimination across environments in peanut. Crop Sci., 34: 98-103.
Direct Link - Nageswara Rao, R.C., M. Udayakumar, G.D. Farquhar, H.S. Talwar and T.G. Prasad, 1995. Variation in carbon isotope discrimination and its relationship to specific leaf area and ribulose-1, 5-bisphosphate carboxylase content in groundnut genotypes. Aust. J. Plant Physiol., 22: 545-551.
CrossRefDirect Link - Nageswara Rao, R.C., H.S. Talwar and G.C. Wright, 2001. Rapid assessment of specific leaf area and leaf nitrogen in peanut (Arachis hypogaea L.) using a chlorophyll meter. J. Agron. Crop Sci., 189: 175-182.
CrossRefDirect Link - Nautiyal, P.C., R.C. Nageswara Rao and Y.C. Joshi, 2002. Moisture-deficit-induced changes in leaf-water content, leaf carbon exchange rate and biomass production in groundnut cultivars differing in specific leaf area. Field Crops Res., 74: 67-79.
CrossRefDirect Link - Ommen, O.E., A. Donnelly, S. Vanhoutvin, M. van Oijen and R. Manderscheid, 1999. Chlorophyll content of spring wheat flag leaves grown under elevated CO2 concentrations and other environmental stresses within the ESPACE-wheat project. Eur. J. Agron., 10: 197-203.
CrossRefDirect Link - Richards, R.A., 1996. Defining selection criteria to improve yield under drought. Plant Growth Regul., 20: 157-166.
CrossRefDirect Link - Richardson, A.D., S.P. Duigan and G.P. Berlyn, 2002. An evaluation of noninvasive methods to estimate foliar chlorophyll content. New Phytol., 153: 185-194.
CrossRefDirect Link - Samdur, M.Y., A.L. Singh, R.K. Mathur, P. Manivel, B.M. ChiKani, H.K. Gor and M.A. Khan, 2000. Field evaluation of chlorophyll meter for screening groundnut (Arachis hypogaea L.) genotype tolerant to iron deficiency chlorosis. Curr. Sci., 79: 211-214.
Direct Link - Sheshshayee, M.S., H. Bindumadhava, N.R. Rachaputi, T.G. Prasad, M. Udayakumar, G.C. Wright and S.N. Nigam, 2006. Leaf chlorophyll concentration relates to transpiration efficiency in peanut. Ann. Applied Biol., 148: 7-15.
CrossRefDirect Link - This, D., C. Borries, I. Souyris and B. Teulat, 2000. QTL study of chlorophyll content as a genetic parameter of drought tolerance in barley. Barley Genetics Newsletter, 30: 20-23.
Direct Link - Turner, F.T. and M.F. Jund, 1994. Assessing the nitrogen requirements of rice crops with a chlorophyll meter method. Aust. J. Exp. Agric., 34: 1001-1005.
CrossRefDirect Link - Uddling, J., J. Gelang-Alfredsson, K. Piikki and H. Pleijel, 2007. Evaluating the relationship between leaf chlorophyll concentration and SPAD-502 chlorophyll meter readings. Photosynth. Res., 91: 37-46.
CrossRefPubMedDirect Link - Van der Mescht, A., J.A. de Ronde and F.T. Rossouw, 1999. Chlorophyll fluorescence and chlorophyll content as a measure of drought tolerance in potato. South Afr. J. Sci., 95: 407-412.
Direct Link - Wright, G.C., K.T. Hubick, G.D. Farquhar and R.C. Nageswara Rao, 1993. Genetic and Environmental Variation in Transpiration Efficiency and its Correlation With Carbon Isotope Discrimination and Specific Leaf Area in Peanut. In: Stable Isotope and Carbon-Water Relations, Ehleringer, J.R., A.E. Hall and G.D. Farquhar (Eds.). Academic Press Inc., New York, ISBN: 0-12-233380-2, pp: 247-267.
- Wu, F.B., L.H. Wu and F.H. Xu, 1998. Chlorophyll meter to predict nitrogen sidedress requirement for short-season cotton (Gossypium hirsutum L.). Field Crops Res., 56: 309-314.
CrossRefDirect Link