
M. Issoufou
Institut des Regions Arides, 4119 Medenine, Tunisia
M. Zaman Allah
SUPAGRO-INRA, Centre International d`Etudes Superieures en Sciences Agronomiques, F 34060 Montpellier Cedex 1, France
A. Ferchichi
Institut des Regions Arides, 4119 Medenine, Tunisia
E. Ferjani
Physiologie et Genetique des Plantes a Intret Agronomique, Faculte des Sciences de Bizerte, 7021 Jarzouna, Tunisia
Asian Journal of Plant Sciences
Year: 2008 | Volume: 7 | Issue: 1 | Page No.: 95-99







ABSTRACT
Buffel grass (Cenchrus ciliaris) growth, biomass allocation to root and shoot, water use efficiency and photosynthetic activity were measured in response to two defoliation heights (5 and 3 cm). Results showed that plants clipped at 5 cm showed a significant increase of diameter and water use efficiency as well as higher leaf water potential with a subsequent increase of stomatal conductance and photosynthetic activity. On the other hand, plants clipped at 3 cm presented an increase in photosynthetic activity parameters but a significant decrease in all growth parameters and water use efficiency.
PDF Abstract XML References Citation
How to cite this article
M. Issoufou, M. Zaman Allah, A. Ferchichi and E. Ferjani, 2008. Clipping Effects on the Growth Variation, Water Use Efficiency and Photosynthetic Activity in Buffel Grass (Cenchrus ciliaris L.) Poaceae
. Asian Journal of Plant Sciences, 7: 95-99.
DOI: 10.3923/ajps.2008.95.99
URL: https://scialert.net/abstract/?doi=ajps.2008.95.99
DOI: 10.3923/ajps.2008.95.99
URL: https://scialert.net/abstract/?doi=ajps.2008.95.99
INTRODUCTION
Bufflel grass (Cenchrus ciliaris syn Pennisetum ciliare) is a C4 bunch grass native to dry areas of the African continent, West Asia and India and has been widely introduced in arid and semi-arid regions of the world (MSeddi et al., 2002). The most drought tolerant of the commonly sown grasses, occurs naturally in areas with average annual rainfall from as low as 100 mm. The species has a larger and deeper root system capable of providing effective soil erosion control in arid and semi-arid regions (Harwood et al., 1999; Gyasi-Agyei et al., 2005). C. ciliaris is typically propagated by seeds that can remain dormant for a considerable period of time before reaching maturity (Burquez-Montijo et al., 2002). In Tunisia, the species is scarce and growth is stunted, because it is selectively overgrazed (Chaieb et al., 1996). Studies on C. ciliaris have concentrated on estimating the nutritional value and preference by animals through intake, diets, crude protein and in vitro organic matter digestibility analysis (Ramirez et al., 1997; Mero and Udén, 1998; Ronquillo et al., 1998; Foroughbackhch et al., 2001). Other studies focused on nitrogen use efficiency (Rudmanna et al., 2001) and on the variation and the relationships of several seed and spike-related traits of C. ciliaris (M`Seddi et al., 2002).
The effects of defoliation on plant growth are complex (Painter and Belsky, 1993): sometimes inhibitory, nil or stimulatory (Oesterheld and McNaughton, 1988, 1991; Georgiadis et al., 1989; Schmid et al., 1990). Understanding the impacts of defoliation height on the performance of grasses is therefore important in the management of grazing areas since animals grazed the plants at different heights (Floret and Pontanier, 1982; Laca et al., 1992). It is especially important in arid and semi-arid environments where, apart from water scarcity, the levels at which grasses are grazed have a major influence on their persistence in the environment.
This study was undertaken to estimate the effect of defoliation height (3 and 5 cm above ground level) on the performance of buffel grass in growth, biomass allocation to shoot and root, water use efficiency and photosynthetic activity.
MATERIALS AND METHODS
This study was carried out at the Institut des Régions Arides de Médenine. The seeds of C. ciliaris were collected during spring 2003 in the protected national park of Bouhedma in southern Tunisia and sown in cylindrical plastic pots (top diameter: 20 cm, height: 18 cm, down diameter: 17 cm) with three drainage holes and containing 4 kg of collected soil: conductivity 5.6 mS cm-1, pH 7.5, total limestone 8%, active limestone 2.5%, gypsum 0.4% and organic mater 0.7%. The seeds were sown to obtain three plants per pot under glasshouse conditions with day/night temperatures of 25/30 °C throughout the experimentation (180 days after sowing). Before sowing, the pots were brought to field capacity. Fifteen Days After Sowing (DAS) and during 6 months, water was supplied at the rate of 20 mm each 15 days corresponding to a total amount of water of 240 mm to ensure adequate soil moisture.
Defoliation treatments: All the plants were submitted to three defoliation treatments: control (C) not cut, C3 (cut at 3 cm above soil surface in the pot) and C5 (cut at 5 cm above soil surface in the pot). The cuts were realized with scissors four times. The first three cuts were realized at 63, 84 and 105 DAS, the last cut occurred at 180 DAS.
Plant growth, Water Use Efficiency (WUE) and photosynthetic activity measurements: Before each cut, plant height and diameter were measured. After each cut, shoots were placed in an oven for 24 h at 105 °C and their dry weight was measured. At the end of the experiment (180 DAS) roots were separated from shoots and both shoots and roots dry weights of all the plants were measured. Water use efficiency was calculated as follows:
Photosynthetic activity was measured from 63 to 105 DAS. Before defoliation, leaf water potential (ψ), stomatal conductance, transpiration rate and photosynthesis rate were measured on six randomly chosen leaves of each clipping treatment using LCi Portable Photosynthesis System (ADC BioScientific CO., UK).
Statistical analysis: Data were analyzed by one-way analysis of variance procedure (ANOVA) using SAS (9.0 for Windows). Subsequent comparison of means was performed using the Duncan`s multiple range test at 5%.
RESULTS AND DISCUSSION
Plant growth and water use efficiency: Defoliation affected C. ciliaris biomass production depending on the cutting height. As compared to the control plants, those defoliated at 5 cm were not affected in shoot biomass but showed a significant decrease of 24.74% in root biomass (Table 1). This indicated that when plants were defoliated to 5 cm, resources were allocated to the shoot over roots for shoot compensation. The ability of 5 cm defoliated plants to compensate shoot biomass resulting in an equal shoot dry mass with the control seemed not to be associated with recovering in plant height but with an increase in plant diameter (Table 1). The increased plant diameter may result from tillering stimulated by the removal of apical dominance after defoliation (Manske, 1994, 2003; Briske and Richards, 1995) but most of the additional tillers remained vegetative and the effect of defoliation on vegetative biomass was generally not significant (Gutman et al., 2001). Compensatory growth following defoliation has long been known in plants but may depend on species, water and nutrients availability as well as severity of grazing (McNaughton, 1984; Gutman et al., 2001). However, 5 cm defoliated plants were not able to completely recover the total biomass and showed a decrease in the root to shoot ratio.
Plants defoliated at 3 cm showed a decrease in all plant growth parameters in a higher way than did defoliation at 5 cm. As compared to the control, defoliation at 3 cm height decreased total biomass by 51%, shoot biomass by 34% and root biomass by 78%, suggesting that roots were more sensitive than shoot when plants were clipped to 3 cm. Sensitivity of root over shoot following defoliation was also observed at 5 cm cut. Thus, defoliation most likely affects more the roots than shoot in buffel grass whatever the clipping height.
Many Pot and field studies have reported a superficial root system development and a reduction of root biomass as well as a decrease of root length after defoliation (McNaughton et al., 1983; Mawdsley and Bardgett, 1997; Manske, 1999, 2000). Chaieb et al. (1996) reported a decrease in root and shoot biomass as well as a reduction of root density, distribution and rooting depth when buffel grass is cut at ground level and under high and moderate cutting frequency.
This may explain the decrease of about 68% in root to shoot ratio after 3 cm defoliation.
In addition, plants cut at 3 cm do not recover and were not able to produce at their potential level; the
| Table 1: | Shoot (S), root (R) and total (T) plant dry weight (DW g plant-1), Root to Shoot Ratio (RSR) and Water Use Efficiency (WUE) of plants subjected or not to cutting at 5 cm (C5) and 3 cm (C3) in Bufflel grass (C. ciliaris) grown under glasshouse conditions |
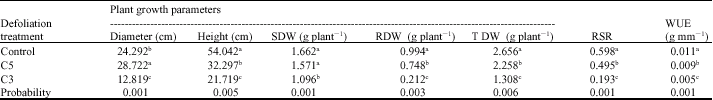 | |
| In columns, means with the same letter(s) are not significantly different | |
compensatory growth was therefore not observed. Present results are in agreement with those obtained for various species (Stockdale, 1992; Virkajärvi, 2003). High defoliation (28% shoot mass or higher than 25% leaf area removed) reduce the root to shoot ratio relative to moderate or non-defoliated Heterotheca subaxillaris plants (Mihaliak and Lincoln, 1989). Clipping to 4 cm reduced both the biomass of root and leaf base of Lolium perenne and Poa trivialis compared with those clipped to 8 cm (Thornton and Millard, 1996). For timothy (Phleum pratense) and meadow fescue (Festuca pratensis) the effect of three defoliation heights, 3, 6 and 9 cm increased the regrowth rates by 27% and the cumulative regrowth dry matter yield increased by 29% of both species by increasing the cutting height from 3- 9 cm (Virkajärvi, 2003). A four weeks defoliation interval of an irrigated Persian clover (Trifolium resupinatum) was more productive when defoliated to 4 cm above ground level than ground level defoliation (Stockdale, 1992).
Water use efficiency expressed as the ratio of total plant biomass to total water received increased with defoliating at 5 cm but decrease with 3 cm clipping level as compared to the control (Table 1). The low WUE at 3 cm defoliation can be attributed to the reduced total biomass production.
Photosynthetic activity: Photosynthetic parameters are presented in Fig. 1(A-D). As defoliation started at 63 DAS, initial leaf water potential (ψ) values were -1.94 MPa for all the plants (Fig. 1A). During all the experimentation, from 63 to 105 DAS, ψ values vary from -1.94 to -2.61 Mpa for the control, to -2.25 MPa for 3 cm defoliated plants and to -1.85 MPa for plants cut to 5 cm. ψ significantly increased with defoliation height at both 84 and 105 DAS. Similar results were obtained with various species (Wolf and Parrish, 1982; Noitsakis and Berger, 1984; Lazaridou, 2001). The low ψ values of the control can be attributed to faster maturation (Link et al., 1990). In contrast, defoliation maintained the plants at the vegetative stage and this effect was accompanied by an increase in ψ. The water potential differences are probably in the same order as the relative water content (Kappen et al., 1994). High ψ values in defoliated plants induced significantly higher values of stomatal conductance (Fig. 1B) and consequently higher transpiration rates at both 84 and 105 DAS (Fig. 1C). The ability of the defoliated plants to maintain an important stomatal conductance is accompanied by a higher photosynthesis rate (Fig. 1D) indicating a higher CO2 uptake. New leaves emerging after defoliation have been found to have greater efficiency for CO2 assimilation
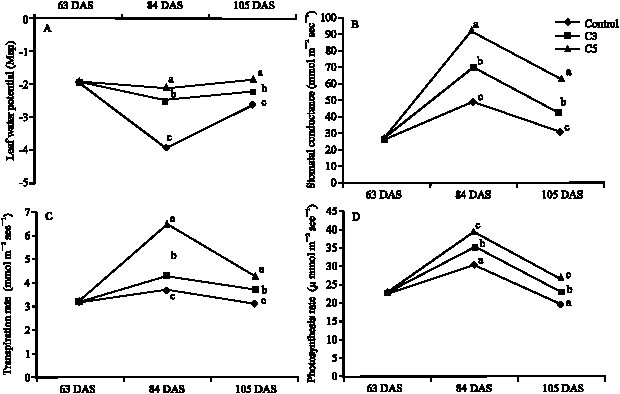 | |
| Fig. 1: | Effects of defoliation height on leaf water potential (A), stomatal conductance (B), transpiration rate (C) and Photosynthesis rate (D). At each corresponding DAS, means indicated by the same letter(s) are not significantly different at 5% |
(Khan and Lone, 2005). Despite the reduction of photosynthetic surfaces by defoliation, clipped plants showed higher photosynthetic capacity. The increase in the photosynthetic parameters of plants defoliated at 84 DAS and 105 DAS was probably due to higher photosynthates requirement of leaves following defoliation for growth and development.
Grazing-tolerant species often respond to defoliation by increasing photosynthetic rates, allocating resources to photosynthetic tissue over roots (Caldwell et al., 1981; McNaughton, 1984; Coughenour et al., 1985). On the other hand the clipping interval and the decrease in root biomass which in turn reduce soil water extraction capacity of 3 cm defoliation plant may explain to reduction to produce at their potential level and therefore can not recover. These results are relevant in the restoration of degraded pasture land based on species sowing methods in arid region.
REFERENCES
- Burquez-Montijo, A., M. Miller and A. Martinez-Yrizar, 2002. Mexican Grasslands, Thornshrub and the Transformation of the Sonoran Desert by Invasive Exotic Buffelgrass (Pennisetum ciliare). In: Invasive Exotic Species in the Sonoran Region, Tellman, B. (Ed.). University of Arizona Press, Tucson, pp: 126-146.
- Caldwell, M., M. Richards, J.H.D.A. Johnson, R.S. Nowak and R.S. Dzuree, 1981. Coping with herbivory: Photosynthetic capacity and resource allocation in 2 semiarid Agropyron bunch grasses. Oecologia, 50: 14-24.
CrossRef - Chaieb, M., B. Henchi and M. Boukhris, 1996. Impact of clipping on root systems of 3 grasses species in Tunisia. J. Range Manage., 49: 336-339.
Direct Link - Coughenour, M.B., S.J. McNaughton and L.L. Wallace, 1985. Responses of African tall-grass (Hyparrhenia filipendula) to defoliation and limitations of water and nitrogen. Oecologia, 68: 80-86.
CrossRef - Foroughbackch, R., R.G. Ramirez, L. Hauad, J. Alba-Avila, C.G. Garcia-Castillo and M. Espinosa-Vazquez, 2001. Dry matter, crude protein and cell wall digestion of total plant, leaves and stems in llano buffelgrass (Cenchrus ciliaris). J. Applied Anim. Res., 20: 181-188.
CrossRefDirect Link - Georgiadis, N.J., R.W. Ruess, S.J. McNaughton and D. Western, 1989. Ecological conditions that determine when grazing stimulates grass production. Oecologia, 81: 316-322.
CrossRef - Gutman, M., I. Noy-Meir, D. Pluda, N.A. Seligman, S. Rothman and M. Sternberg, 2001. Biomass partitioning following defoliation of annual and perennial Mediterranean grasses. Conserv. Ecol., 5: 1-1.
Direct Link - Harwood, M.R., J.B. Hacker and J.J. Mott, 1999. Field evaluation of seven grasses for use in the revegetation of lands disturbed by coal mining in central Queensland. Aust. J. Exp. Agric., 39: 307-316.
Direct Link - Kappen, L., G. Schultz and R. Vanselow, 1994. Direct observations of stomatal movements. Ecol. Stud., 100: 231-246.
Direct Link - Khan, N.A. and P.M. Lone, 2005. Effects of early and late season defoliation on photosynthesis, growth and yield of mustard (Brassica juncea L.). Brazilian J. Plant Physiol., 17: 181-186.
CrossRefDirect Link - Laca, E.A., E.D. Ungar, N.G. Seligman and M.W. Demment, 1992. Effects of sward height and bulk density on bite dimensions of cattle grazing homogeneous swards. Grass Forage Sci., 47: 91-102.
CrossRef - Link, S.O., G.W. Gee and J.L. Downs, 1990. The effect of water stress on phenological and ecophysiological characteristics of cheatgrass and Sandberg's bluegrass. J. R. Manage., 43: 506-513.
Direct Link - Mawdsley, J.L. and R.D. Bardgett, 1997. Continuous defoliation of perennial ryegrass (Lolium perenne) and white clover (Trifolium repens) and associated changes in the composition and activity of the microbial population of an upland grassland soil. Biol. Fertil. Soils, 24: 52-58.
CrossRefDirect Link - McNaughton, S.J., L.L. Wallace and M.B. Coughenour, 1983. Plant adaptation in an ecosystem context: Effects of defoliation, nitrogen and water on growth of an African C4 sedge. Ecology, 64: 307-318.
CrossRefDirect Link - McNaughton, S.J., 1984. Grazing lawns: Animals in herds, plant form and coevolution. Am. Nat., 124: 863-886.
CrossRefDirect Link - Mero, R.N. and P. Uden, 1998. Promising tropical grasses and legumes as feed resources in Central Tanzania V. Effect of supplementing Cenchrus ciliaris hay with leaves from four legumes on intake and digestibility by growing Mpwapwa bulls. Anim. Feed Sci. Technol., 70: 111-122.
Direct Link - Mihaliak, C.A. and D.E. Lincoln, 1989. Plant biomass partitioning and chemical defense: Response to defoliation and nitrate limitation. Oecologia, 80: 122-126.
CrossRef - M'Seddi, K., M. Visser, M. Neffati, D. Reheul and M. Chaieb, 2002. Seed and spike traits from remnant populations of Cenchrus ciliaris L. in South Tunisia: High distinctiveness, no ecotypes. J. Arid Environ., 50: 309-3254.
CrossRef - Noitsakis, B. and A. Berger, 1984. Relations hydriques chez Dactylis glomerata et Dichanthium ischaeum cultivés sous deux régimes hydriques contrastes. Acta Oecol. Plant, 5: 75-88.
Direct Link - Oesterheld, M. and S.J. McNaughton, 1988. Intraspecific variation in the response of Themeda triandra to defoliation: The effect of time of recovery and growth rates on compensatory growth. Oecologia, 77: 181-186.
CrossRef - Oesterheld, M. and S.J. McNaughton, 1991. Effect of stress and time for recovery on the amount of compensatory growth after grazing. Oecologia, 85: 305-313.
CrossRef - Painter, E.L. and A.J. Belsky, 1993. Application of herbivore optimization theory to rangelands of the western United States. Ecol. Applied, 3: 2-9.
Direct Link - Ramirez, R.G., J.B. Quintanilla and J. Aranda, 1997. White-tailed deer food habits in Northeastern Mexico. Small Ruminant Res., 25: 141-146.
CrossRef - Ronquillo, M.G., M. Fondevila, A.B. Urdaneta and Y. Newman, 1998. In vitro gas production from buffel grass (Cenchrus ciliaris L.) fermentation in relation to the cutting interval, the level of nitrogen fertilisation and the season of growth. Anim. Feed Sci. Technol., 72: 19-32.
CrossRefDirect Link - Rudmanna, S.G., P.J. Milhamb and J.P. Conroy, 2001. Influence of High CO2 Partial Pressure on Nitrogen Use Efficiency of the C4 Grasses Panicum coloratum and Cenchrus ciliaris. Ann. Bot., 88: 571-577.
CrossRefDirect Link - Schmid, B., S.L. Miao and F.A. Bazzaz, 1990. Effects of simulated root herbivory and fertilizer application on growth and biomass allocation in the clonal perennial Solidago canadensis. Oecologia, 84: 9-15.
CrossRef - Stockdale, C.R., 1992. Effects of frequency and height of defoliation on the production of a Persian clover-annual ryegrass sward. Aust. J. Exp. Agric., 32: 339-344.
CrossRef - Thornton, B. and P. Millard, 1996. Effects of severity of defoliation on root functioning in grasses. J. R. Manage., 49: 443-447.
Direct Link - Virkajärvi, P., 2003. Effects of defoliation height on regrowth of timothy and meadow fescue in the generative and vegetative phases of growth. Agric. F. Sci., 12: 177-193.
Direct Link - Wolf, D.D. and D.J. Parrish, 1982. Short-term growth responses of tall fescue to changes in soil water potential and to defoliation. Crop Sci., 22: 996-997.
Direct Link