
D. Nava-Arenas
Departamento de Graduados e Investigacion en Alimentos, Escuela Nacional de Ciencias Biologicas, Instituto Politecnico Nacional, Carpio y Plan de Ayala, CP 11340 Mexico, DF, Mexico
A. Jimenez-Aparicio
Departamento de Biotecnologia, Centro de Desarrollo de Productos Bioticos, Instituto Politecnico Nacional, Yautepec, Morelos, Mexico
H. Hernandez-Sanchez
Departamento de Graduados e Investigacion en Alimentos, Escuela Nacional de Ciencias Biologicas, Instituto Politecnico Nacional, Carpio y Plan de Ayala, CP 11340 Mexico, DF, Mexico
Asian Journal of Plant Sciences
Year: 2008 | Volume: 7 | Issue: 7 | Page No.: 682-686







ABSTRACT
A Taguchi L9 orthogonal array methodology was used to optimize the conditions for maximal germination percentage in blue corn grains. The factors studied included steeping time, ratio of steeping water to corn grain, germination temperature and time. The optimal conditions included a steeping time of 24 h, an amount of 100 g of grain steeped in 200 mL of water and a germination time of 96 h at 25°C for a predicted germination of 90.6%. Under these conditions, 94% of the grains germinated reaching a moisture content of 43%. An increase in the range of 154 to 160% for total phenolics and 43% for anthocyanins was obtained after 96 h of germination. The 1,1-diphenyl-2-picrylhydrazyl radical (DPPH) antioxidant activity increased slightly during the germination process. In conclusion, the process of germination was beneficial due to the increase in antioxidant compounds in the blue corn grain.
PDF Abstract XML References Citation
How to cite this article
D. Nava-Arenas, A. Jimenez-Aparicio and H. Hernandez-Sanchez, 2008. Optimization of Germination Conditions of Blue Corn (Zea mays L.) by Taguchi Orthogonal Array Methodology. Asian Journal of Plant Sciences, 7: 682-686.
DOI: 10.3923/ajps.2008.682.686
URL: https://scialert.net/abstract/?doi=ajps.2008.682.686
DOI: 10.3923/ajps.2008.682.686
URL: https://scialert.net/abstract/?doi=ajps.2008.682.686
INTRODUCTION
Germination is the first step in the production of a native white or yellow corn beer called tesgüino which is consumed by several ethnic and mestizo groups in Mexico. Tesgüino is mainly consumed during ritualistic ceremonies by the Tarahumara and some other native tribes in Chihuahua but some light versions of the product are sold as refreshing beverages in the streets of several towns in Northern Mexico (Quintero-Ramírez et al., 1999; Blandino et al., 2003). For the preparation of tesgüino, dark-germinated corn is ground to obtain a dough which is boiled in water. Liquid portions are transferred to a clay pot and a catalyst is added. This mixture is allowed to ferment for two or three days to obtain the final product. The bacteria and yeasts isolated from tesgüino include mainly Lactobacillus, Streptococcus, Leuconostoc, Pediococcus and Saccharomyces (Quintero-Ramírez et al., 1999).
No studies are reported about the use of blue corn, which has a high content of anthocyanins, in the elaboration of tesgüino. Anthocyanins are hydrosoluble vegetable pigments responsible for the red, blue and purple color of many fruits, flowers and seeds. They occur mainly as glycosides (Wu et al., 2006). Cyanidin 3-glucoside has been identified as the major anthocyanin (75%) in Mexican blue corn from Toluca, which contains 321 mg kg-1 of these compounds and 451 mg kg-1 of total polyphenolic compounds (Del Pozo-Insfran et al., 2006). These compounds may have an important role in scavenging the active oxygen species which are generated during seed germination (Bailly, 2004) and have been reported to be important in the prevention of obesity and in the amelioration of hyperglycemia in mice (Tsuda et al., 2003). The use of this particular cereal could produce a tesgüino with a good content of antioxidant compounds.
Many cereals can be satisfactory malted, but corn is rarely used for this purpose because its fat becomes rancid during storage (Hough, 1991). However, it has been reported that some yellow corn varieties had good malting performance (Eneje et al., 2004) and no problems have been detected when the corn malt is used immediately after its preparation for the elaboration of beer-like products like chicha or tesgüino (Quintero-Ramírez et al., 1999). The main purpose of malting is the modification of the seed storage starch and protein reserves into assimilable materials for adequate yeast growth and fermentation (Ogbonna et al., 2003).
Germination is a process that has been optimized for some cereals (Capanzana and Buckle, 1997). Germination starts once a seed takes up water. Seed steeping is a known strategy to increase the uniformity of seed germination, but in the case of corn and rice, water-steeping also accelerates the germination process (Horii et al., 2007). Steeping is normally completed in two days and then the grains are drained and spread in germination trays or incubators. There are studies indicating that the steeping and germination time and temperature affect the malting performance and fermentability of grains (Eneje et al., 2004; Balogun et al., 2006).
Most published optimization procedures involve response surface methodology, however, when the number of variables is high, the number of experimental runs is also very high. Reports are available on the use of Taguchi orthogonal matrix method in biotechnology, since this method examine the effects of several process variables and identify the factors which have major effects on the process using just a few experiments (Seyedeh et al., 2007).
This study reports the manipulation of steeping time, amount of steeping water and germination time and temperature for the optimization of germination percentage of blue corn using the Taguchi method. Since germination can modify the phenolic composition of seeds (López-Amorós et al., 2006), the changes in phenolic compounds and anthocyanins during germination of blue corn were also studied.
MATERIALS AND METHODS
The study was conducted in 2007 at the Escuela Nacional de Ciencias Biológicas of the Instituto Politécnico Nacional in Mexico City, DF, Mexico.
Analytical methods: The blue corn grain was purchased in the town of San Mateo Tepopula, State of Mexico (near Mexico City) and was analyzed for moisture (44-16), fat (30-10), protein (46-10), fiber (32-10), ash (08-01) and carbohydrates (80-60) by the approved methods of the AACC (AACC, 2000). The grains were physically and biologically characterized with the following parameters: hectoliter weight (Salinas-Moreno et al., 2007), seed viability by the tetrazolium test (Rao et al., 2006), germinative energy (Langrell and Edney, 2006), germinative capacity (Langrell and Edney, 2006) and amount of impurities (Piedade et al., 2002). Phenolics were extracted with acidic methanol and measured with the Folin-Ciocalteau technique (Velioglu et al., 1998). Anthocyanins were extracted with acidic ethanol and its absorbance measured at 535 nm (Abdel-Aal and Hucl, 1999).
| Table 1: | L9 orthogonal array for the germination process |
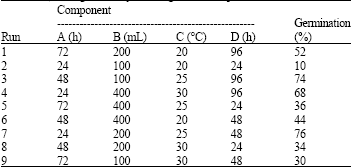 | |
| A: Steeping time; B: Amount of steeping water per 100 g of corn grain; C: Germination temperature; D: Germination time | |
The content of these compounds was also evaluated in white corn from the same region. The antioxidant activity was measured as DPPH (1,1-diphenyl-2-picrylhydrazyl)-radical scavenging activity. Samples (0.2 g fresh weight) were ground in a mortar at 4°C in 2.0 mL of absolute ethanol. One-half milliliter of the solution was mixed with 0.25 mL of 0.5 mM DPPH in ethanol solution and 0.5 mL of 100 mM acetate buffer (pH 5.5). The absorbance of the mixture was measured at 517 nm after 30 min (Kang and Saltveit, 2002) and compared with the absorbance of a solution without sample. The inhibition (%) of the color formation was reported. All the results are shown as the average of three determinations.
Optimization of the germination conditions using the Taguchi L9 orthogonal array methodology: The design for the L9 orthogonal array was developed and analyzed using the Design Expert 7.0.3 software (Stat-Ease Inc., Minneapolis, MN 55413). The factors studied included steeping time (24, 48 and 72 h), amount of steeping water (100, 200 and 400 mL water) used per 100 g of corn grain, germination temperature (20, 25 and 30°C) and time (24, 48 and 96 h). Table 1 shows the dark-germination conditions and the L9 orthogonal array used in this study. The seeds were steeped in water and then placed into Petri dishes containing two discs of Whatman No. 1 filter paper which were moistened with distilled water to make certain that the moisture was adequate for germination (Koduru et al., 2006). Controlled temperature chambers were used for seed incubation at the different temperatures. Three replicates of 100 seeds were used for each assay.
RESULTS AND DISCUSSION
Blue corn composition and properties: The results of the proximal analysis of the blue corn grain are shown in Table 2. These results are very similar to those previously reported for white and yellow corn (Haard, 1999; Eneje et al., 2004).
| Table 2: | Blue corn grain composition |
 | |
| Table 3: | Some physical and germinative characteristics of the blue corn seeds |
 | |
Some other characteristics of the corn seeds are shown in Table 3. The hectoliter weight falls in the normal range for corn (61.0-73.6 kg HL-1) reported by Greenway et al. (1977). The grain viability was relatively high. A viable seed is one, which is capable of germinating under the proper circumstances, so 92% viability could be correlated with a high probability of germination. The germinative energy and capacity for the blue corn grains (70.4 and 93%, respectively) were low compared with the values reported for white (95 and 97%) and yellow (97 and 99%) corns (Eneje et al., 2004). More studies about these parameters using different varieties of blue corn from several places would be necessary to conclude adequately about the germinative properties of this pigmented seed.
The amount of anthocyanins was, as expected, considerably higher in blue than in white corn. The concentration of anthocyanins in the blue corn used in this study (540 mg kg-1 corn) was higher than the content reported by Del Pozo-Insfran et al. (2006) for American (307 mg kg-1) and Mexican (321 mg kg-1) blue corns. The total polyphenolic content of the white corn (1741 mg kg-1) was similar to that reported by Adom and Liu (2002) for yellow corn (1758 mg kg-1), however, the concentration of polyphenols in the blue corn in this study (1528 mg kg-1) was considerably lower than the content indicated by Del Pozo-Insfran et al. (2006) for a Mexican white corn from Monterrey (4899 mg kg-1). The antioxidant activity for the blue corn allowed a 52% of inhibition of the DPPH color formation.
Optimization of the germination process: The optimal conditions predicted by the software included a steeping time of 24 h, an amount of 100 g of grain steeped in 200 mL of water and a germination time of 96 h at 25°C for a predicted maximal grain germination of 90.6% (Table 1). The four factors had an equal statistical importance level (25%). When these conditions were tested experimentally, 94% of the grains germinated reaching a final moisture content of 43%. The 2:1 water/grain ratio is the same proportion recommended for malting sorghum (Ogbonna et al., 2003).
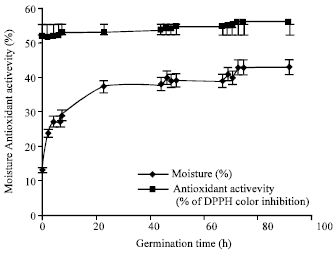 | |
| Fig. 1: | Time courses of moisture content (%) and antioxidant activity (% of DPPH color inhibition) during the germination process of blue corn at 25°C |
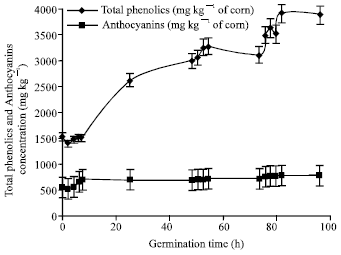 | |
| Fig. 2: | Time courses of total phenolic content (mg kg-1) and anthocyanins content (mg kg-1) during the germination process of blue corn at 25°C |
After the 24 h steeping period, moisture increased from 13.1 to 37.41%, phenolics increased from 1528 to 2600 mg kg-1 of corn (expressed as ferulic acid) and anthocyanins increased from 540 to 678 mg kg-1 of corn (expressed as cyanidine-3-glucoside). The grains were then germinated for 96 h. Moisture, phenolics and anthocyanins increased to important final contents of 43.01%, 3800 and 775 mg kg-1, respectively (Fig. 1, 2). The antioxidant activity increased to 56% of inhibition of the DPPH color formation due to an increase in the content of phenolics. These results confirm the findings of Randhir and Shetty (2005) about the developmental stimulation of total phenolics and their antioxidant activity in dark germinated sweet corn. They found that the phenylpropanoid pathway was stimulated during dark-germination of corn and consequently the total phenolic content increased to a value (4000 mg kg-1) similar to the one found in this study for blue corn. They also noticed an increase in antioxidant activity to 81% DPPH inhibition after 8 days of germination. Similar increases in phenolics and antioxidant activity have also been observed during the germination of wheat, fava bean and Lupinus angustifolius (Yang et al., 2001; Shetty et al., 2003; Fernández-Orozco et al., 2006). However, in the case of sorghum, germination did not affect the content of total phenolic compounds (Dicko et al., 2005).
The increases in moisture, phenolic compounds and anthocyanins during the germination process could be fitted to the following equations:
 |
where, t is germination time in hours and R2 is the determination coefficient of the equation fitting the data in Fig. 1 and 2.
It can be concluded that the content of anthocyanins is, as expected, considerably higher in the blue corn with respect to the white grain. However, the content of total phenolic compounds is slightly higher in the white corn. The germination process was shown to be beneficial due to the increase in anthocyanins, total phenolics and antioxidant activity in the blue corn malt. The optimization of the germination process can ensure a maximal proportion of sprouted grains and with it a higher yield of blue tesgüino. This research provides a new tool for the optimization of processes such as germination.
ACKNOWLEDGMENT
The Research was supported by the Instituto Politécnico Nacional through the grant CGPI 20060568 and by COFAA-IPN fellowships.
REFERENCES
- Abdel-Aal, E.S.M. and P. Hucl, 1999. A rapid method for quantifying total anthocyanins in blue aleurone and purple pericarp wheats. Cereal Chem., 76: 350-354.
CrossRefDirect Link - Adom, K.K. and R.H. Liu, 2002. Antioxidant activity of grains. J. Agric. Food Chem., 50: 6182-6187.
CrossRefDirect Link - Bailly, C., 2004. Active oxygen species and antioxidants in seed biology. Seed Sci. Res., 14: 93-107.
CrossRefDirect Link - Balogun, R.O., S.H. Bird and J.B. Rowe, 2006. Germination temperature and time affect in vitro fermentability of sorghum grain. Animal Feed Sci. Technol., 127: 125-132.
Direct Link - Blandino, A., M.E. Al-Aseeri, S.S. Pandiella, D. Cantero and C. Webb, 2003. Cereal-based fermented foods and beverages. Food Res. Int., 36: 527-543.
CrossRefDirect Link - Capanzana, M.V. and K.A. Buckle, 1997. Optimisation of germination conditions by response surface methodology of a high amylase rice (Oriza sativa) cultivar. Lebensm. Wiss. Technol., 30: 155-163.
CrossRef - Del Pozo-Insfran, D., C.H. Brenes, S.O. Serna and S.T. Talcott, 2006. Polyphenolic and antioxidant content of white and blue corn (Zea mays L.) products. Food Res. Int., 39: 696-703.
Direct Link - Dicko, M.H., H. Gruppen, A.S. Traore, W.J.H. Van Berkel and A.G.J. Voragen, 2005. Evaluation of the effect of germination on phenolic compounds and antioxidant activities in sorghum varieties. J. Agric. Food Chem., 53: 2581-2588.
Direct Link - Eneje, L.O., E.O. Ogu, C.U. Aloh, F.J.C. Odibo, R.C. Agu and G.H. Palmer, 2004. Effect of steeping and germination time on malting performance of Nigerian white and yellow corn varieties. Process Biochem., 39: 1013-1016.
CrossRefDirect Link - Fernandez-Orozco, R., M.K. Piskula, H. Zielinski, H. Kozlowska, J. Frias and C. Vidal-Valverde, 2006. Germination as a process to improve the antioxidant capacity of Lupinus angustifolius L. var. Zapaton. Eur. Food Res. Technol., 223: 495-502.
CrossRefDirect Link - Greenway, W.T., C.A. Watson and G. Davis, 1977. Factors for converting Bushel weight to hectoliter weight for six cereal grains, flax, and soybeans. Cereal Chem., 54: 373-378.
Direct Link - Horii, A., P. McCue and K. Shetty, 2007. Enhancement of seed vigour following insecticide and phenolic treatment. Bioresour. Technol., 98: 623-632.
Direct Link - Kang, H. and M. Saltveit, 2002. Antioxidant enzymes and DPPH-radical scavenging activity in chilled and heat-shocked rice seedling radicles. J. Agric. Food Chem., 50: 513-518.
Direct Link - Koduru, S., D.S. Grierson and A.J. Afolayan, 2006. Effect of high temperatures on seed germination of Solanum aculeastrum. Asian J. Plant Sci., 5: 353-356.
CrossRefDirect Link - Lopez-Amoros, M.L., T. Hernandez and I. Estrella, 2006. Effect of germination on legume phenolic compounds and their antioxidant activity. J. Food Compos. Anal., 19: 277-283.
CrossRefDirect Link - Ogbonna, A.C., S.K.C. Obi and B.N. Okolo, 2003. Protein modification in malting sorghum. World J. Microbiol. Biotechnol., 19: 495-503.
Direct Link - Piedade, F.S., H. Fonseca, E.M. Gloria, M.A. Calori, S.M. Stefano and D. Barbin, 2002. Distribution of aflatoxins in corn fractions visually segregated for defects. ras. J. Microbiol., 33: 250-254.
Direct Link - Randhir, R. and K. Shetty, 2005. Developmental stimulation of total phenolics and related antioxidant activity in light- and dark-germinated corn by natural elicitors. Process Biochem., 40: 1721-1732.
Direct Link - Rao, N.K., J. Hanson, M.E. Dulloo, K. Ghosh, D. Nowell and M. Larinde, 2006. Manual of Seed Handling in Genebanks. 1st Edn., Handbooks for Genebanks No. 8., Rome, Italy, ISBN: 9789290437406, Pages: 147.
Direct Link - Salinas-Moreno, Y., J.J. López-Reynoso, G.B. González-Flores and G. Vázquez-Carrillo, 2007. Phenolic compounds of maize grain and their relationship with darkening in dough and tortilla. Agrociencia, 41: 295-305.
Direct Link - Seyedeh, F.G.O., T. Fatemeh, B. Yakhchali and E. Feresshteh, 2007. Enhancement of alkaline protease production by Bacillus clausii using Taguchi experimental design. Afr. J. Biotechnol., 6: 2559-2564.
Direct Link - Shetty, P., M.T. Atallah and K. Shetty, 2003. Stimulation of total phenolics, L-DOPA and antioxidant activity through proline-linked pentose phosphate pathway in response to proline and its analogue in germinating fava beans (Vicia faba). Process Biochem., 38: 1707-1717.
Direct Link - Tsuda, T., F. Horio, K. Uchida, H. Aoki and T. Osawa, 2003. Dietary cyanidin 3-O-ß-D-glucoside-rich purple corn color prevents obesity and ameliorates hyperglycemia in mice. J. Nutr., 133: 2125-2130.
Direct Link - Velioglu, Y.S., G. Mazza, L. Gao and B.D. Oomah, 1998. Antioxidant activity and total phenolics in selected fruits, vegetables and grain products. J. Agric. Food Chem., 46: 4113-4117.
CrossRefDirect Link - Wu, X., G.R. Beecher, J.M. Holden, D.B. Haytowitz, S.E. Gebhardt and R.L. Prior, 2006. Concentrations of anthocyanins in common foods in the United States and estimation of normal consumption. J. Agric. Food Chem., 54: 4069-4075.
CrossRefDirect Link - Yang, F., T.K. Basu and B. Ooraikul, 2001. Studies on germination conditions and antioxidant contents of wheat grain. Int. J. Food Sci. Nutr., 52: 319-330.
CrossRefDirect Link