
P. Prathepha
Rice Genetics Research Unit, Department of Biotechnology, Faculty of Technology, Mahasarakham University, Kham Riang District, Maha Sarakham Province 44150, Thailand
Asian Journal of Plant Sciences
Year: 2008 | Volume: 7 | Issue: 6 | Page No.: 599-602







ABSTRACT
Fragrance is the most important trait among the domesticated characteristics of rice (O. sativa L.). The recessive fragrance gene on chromosome 8 is associated with rice fragrance. The gene for fragrance in a fragrant rice variety shows the presence of a mutation portion (i.e., an eight base pair deletion in exon 7). This allele is responsible for rice fragrance. In the present study, 65 self-supplied seed lots of black rice (O. sativa L.) from Thailand and Laos were assessed for purity of the fragrance grain of each sample for this locus using a PCR assay. The results indicate that black rice germplasm were genotyped for homozygous fragrant, heterozygote and non-fragrant homozygous. The data from the 65 samples show that 7.7 and 23% of the samples were heterozygous and non-fragrant homozygous, respectively. Heterozygous individuals, black rice plants that carry both the fragrant allele and non-fragrant allele of the fragrance gene and non-fragrant seeds need to avoid because they are non-fragrant and give rise to a mixture of fragrant and non-fragrant seed lots. Therefore, domestication of black rice in order to maintain grain aroma would require the use of quality black rice seed germplasm.
PDF Abstract XML References Citation
How to cite this article
P. Prathepha, 2008. Evaluation of the Fragrance Gene (fgr) in Self-Supplied Seed
Lots of Black Rice (Oryza sativa L.) from Thailand and Laos. Asian Journal of Plant Sciences, 7: 599-602.
DOI: 10.3923/ajps.2008.599.602
URL: https://scialert.net/abstract/?doi=ajps.2008.599.602
DOI: 10.3923/ajps.2008.599.602
URL: https://scialert.net/abstract/?doi=ajps.2008.599.602
INTRODUCTION
Colored rice, most of which is red or black, is not the most commonly consumed; that distinction belongs to white rice. Colored rice has been considered a health food; black rice pigment fraction has strong preventive effects against atherosclerotic disease or coronary heart disease (Ling et al., 2001, 2002; Xia et al., 2003).
Black rice is popular in Asian countries where it is mixed with white rice prior to cooking to enhance the flavor, color and nutritional value (Yang et al., 2008). Historically, black rice has been reserved for use in festival foods and desserts in Asian countries. Typically, black rice grains are aromatic and because grain fragrance is an important feature of premium-value rice, it commands higher prices in domestic and international markets. In Southeast Asia, especially in Lao PDR and the north and northeastern regions of Thailand, black rice serves as the staple food; Khao Kam is the traditional name of black rice in these regions. Black rice is classified into two categories: grain with purple pigmentation on glumes and various color shades on the pericarp and grain with straw glumes and purple pericarp. Its color can be attributed to anthocyanins (cyaniding 3-glucoside and peonidin 3-glucoside) found in surface cells of the grain (Xia et al., 2006).
Fragrance in rice is a highly valued trait and known to be primarily associated with grain 2-acetyl-1-pyrroline. It has been previously determined that the fragrance gene is located on chromosome 8 that controls the level of aromatic compound 2-acetyl-1-pyrroline (Bradbury et al., 2005a). The structure of the fragrance gene (fgr) comprises 15 exons interrupted by 14 introns. Fragrance is a recessive trait, the alleles from fragrant varieties all showed the presence of mutations (i.e., the 8 bp deletion in exon 7), resulting in a loss of function of the fragrance gene product. Interestingly, the concentration of 2-acetyl-1-pyrroline (2-AP) was high in cooked black rice (Yang et al., 2008). Seed of local rice varieties maintained by farmers are genetically diverse (Saito et al., 2007). Analysis of molecular diversity using molecular techniques allows variation to be evaluated between individual plants, particularly the aromatic character of the black rice of saved farmer seeds, by using a polymerase chain reaction (PCR) assay.
The goal of this study is to genetically characterize the farmers` seed germplasm. The results from this study can be used to enhance the efficiency and effectiveness of black rice germplasm maintenance for the aromatic character, promote farmer awareness of the value of collecting seeds and encourage seed conservation of black rice varieties.
MATERIALS AND METHODS
Sample collection: Seeds of black rice samples from different farmers` self-supplied seed lots in several regions of Thailand and Laos were collected. A total of 65 samples were collected and evaluated. Four monthly collections were made over a period of two years: during March 2006 and 2007 and December 2006 and 2007. The summary information of black rice samples are listed in Table 1.
DNA extraction and polymerase chain reaction: Mature seeds of each seed lot were germinated and grown in pots at a green house at Mahasarakham University. Five seedlings generated from each seed lot were randomly selected and bulked for use in the analysis. Genomic DNA was extracted from the bulked samples of the young leaf according to the protocol of Doyle and Doyle (1987). PCR amplification of the fgr gene was performed using the DNA sequences of oligonucleotide primers (i.e., Os2AP-exon7.1F: 5`-TGCTCCTTTGTCAT CACACC-3` and Os2AP-exon7.1R: 5`-TTTCCACCAAGTTCC AGTGA-3`), which were used previously to amplify the fragrance gene located on chromosome 8. In addition, this DNA marker can be used in breeding for fragrant rice varieties (Shi et al., 2008). The oligonucleotide primers were synthesized by BSU (BioService Unit, National Center for Genetic Engineering and Biotechnology, National Science and Technology Development Agency, Bangkok, Thailand).
The PCR reaction was performed in a 20 μL reaction mixture containing 2 μL of DNA solution, 50 pmol each of the primer pairs, 2.0 mM MgCl2, 2 units Taq polymerase (Promega), 0.1 mM dNTPs. Cycling conditions were 94°C (5 min); then 40 cycles of 94°C (1 min), 60°C (1 min), 72°C (1.5 min) and a final extension of 72°C (5 min). Using these primer pairs, the DNA template from fragrant rice, Khao Dawk Mali 105 and a non-fragrant rice, Chai Nart 1 (CN1), were used as positive and negative controls, respectively, in the experiment for comparison of bands resulting from PCR between fragrant and non-fragrant rice. The PCR products were separated in 4.5% polyacrylamide denaturing gels of 200x125x1 mm (lengthxwidthx thickness). After electrophoresis, the bands were stained with silver-stain. The PCR product of approximately 396 bp obtained from Thai jasmine rice (Khao Dawk Mali 105) was present in every sample with the recessive allele (the 8 bp deletion), whereas the dominant allele gave a product of approximately 404 bp from the Thai non-fragrant rice (CN 1). From the PCR assay, heterozygotes can be discriminated by the presence of both PCR products. The genotypic and allelic frequencies were computed based on Hardy-Weinberg formulations. Goodness-of-fit statistics were calculated for the figure observed compared to values expected using the Hardy-Weinberg equilibrium.
| Table 1: | List of self-supplied seed of black rice (O. sativa L.) of different origins and their PCR-based genotypes of the fragrance gene |
 | |
RESULTS
A PCR assay was used to evaluate 65 self-supplied seed lot plants which derived from seed germination. The assay predicted the genotype of each of the individuals within a seed lot. The different genotypes and the distribution of the allele of the fgr gene in the black rice samples used in the present study are shown in Table 1. Overall, a total number of 45, 5 and 15 seed lots were genotyped for DD (allele D, 8 bp deletion), ND (heterozygote) and NN (allele N, non-deletion), respectively. The data from the 65 combined seed lots show that 31% of the samples were heterozygous and homozygous non-fragrant (Fig 1). The overall allelic frequencies were 0.731 and 0.269 for D and N alleles, respectively. The distribution did differ significantly from that expected under the Hardy-Weinberg equilibrium (Goodness-of-fit χ2 = 42.3, p<0.01) (Table 2). One of the most important causes that may underline the inconsistent results from the Hardy-Weinberg equilibrium is artificial selection of black rice genotype by traditional farmers. The artificial selection causes changes in allele frequencies of the fragrance gene in black rice populations.
| Table 2: | Statistical analysis of fragrance gene (fgr) in the 65 self-supplied seed samples of black rice from Thailand and Laos |
 | |
| Goodness-of-fit testing showed significant difference suggesting that locus of the fgr gene is not conformed to Hardy-Weinberg equilibrium | |
DISCUSSION
A number of sensory methods have been utilized to assist breeders in selecting fragrant rice, but limitations occur when processing large numbers of samples. In addition, these methods are labor intensive, difficult and unreliable (Bradbury et al., 2005b). DNA markers are pieces of DNA that associate the presence or absence of particular traits. Selection for the trait can be undertaken on the basis of molecular techniques. For the fragrance trait in rice, a DNA-based marker situated within the fragrance gene was developed (Bradbury et al., 2005b; Shi et al., 2008). The marker-assisted selection can separate fragrant and non-fragrant rice varieties. An example of the DNA marker approach for screening black rice grain aroma was reported by Bounphanousay et al. (2008). They reported that the fragrant diagnostic band is associated with aromatic character which is indicated by the level of 2-AP in the examined grains. All accessions of their sample black rice showed either homozygous fragrant or homozygous non-fragrant. This means that the seed of black rice used in their report contained a mixture of non-fragrant seed and fragrant seed.
The major finding of the present study was that farmer saved seed lots of black rice had dramatically contaminated (5 out of 65 seed lots) seed of black rice with genotypes non-fragrant and heterozygous.
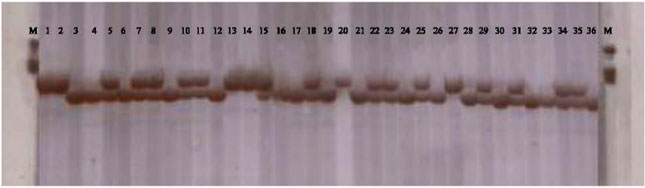 | |
| Fig. 1: | PCR products in a 4.5% polyacrylamide gel showing three genotypes (NN = No-deletion of the 8 base pair; ND = Heterozygote and DD = Deletion of the 8 base pair) of the fgr gene in black rice (O. sativa L.). Lane M shows DNA molecular weight (base pair, bp). Lane, putative samples and their genotypes (respectively) are as follows: NN: 1, 2, 13, 14, 20, 27; ND: 5, 7, 8, 10, 11, 15, 18, 22, 23, 25, 29, 31, 34, 35; DD: 3, 4, 6, 9, 12, 16, 17, 19, 21, 24, 26, 28, 30, 32, 33, 36 |
Heterozygous individuals, black rice plants that carry both the fragrant allele and non-fragrant allele of the fragrance gene, need to be avoided because they are non-fragrant and give rise to a mixture of fragrant and non-fragrant seed lots. Domestication of black rice in order to maintain grain aroma would require the use of quality black rice seed germplasm. The assay can also identify mixtures of fragrant and non-fragrant plants which is useful for pure seed maintenance. Farmer and industry awareness of seed quality can be enhanced by facilitating the development of marketplace demand-driven germplasm improvement. An emphasis on the development of the production of black rice seed quality should become a priority. The small scale farming sector stands to benefit from the development through an increase in productivity, profitability and sustainability.
ACKNOWLEDGMENTS
This study was supported by Mahasarakham University. The author is grateful to Thai and Laos farmers for providing valuable black rice samples and to V. Pilap for his technical assistance.
REFERENCES
- Bradbury, L.M.T., R.J. Henry, Q. Jin, R.F. Reinke and D.L.E. Waters, 2005. A perfect marker for fragrance genotying in rice. Mol. Breed., 16: 279-283.
CrossRef - Bounphanousay, C. P. Jaisil, J. Sanitchon, M. Fitzgerald, N.R.S. Hamilton and J. Sanitchon, 2008. Chemical and molecular characterization of fragrance in black glutinous rice from lao PDR. Asian J. Plant Sci., 7: 1-7.
CrossRefDirect Link - Doyle, J.J. and J.L. Doyle, 1987. A rapid DNA isolation procedure for small quantities of fresh leaf tissue. Phytochem. Bull., 19: 11-15.
Direct Link - Ling, W.H., Q.X. Cheng, J. Ma and T. Wang, 2001. Red and black rice decrease antherosclerotic plaque formation and increase antioxidant status in rabbits. J. Nutr., 131: 1421-1426.
PubMed - Ling, W.H., L.L. Wang and J. Ma, 2002. Supplementation of the black rice outer layer fraction to rabbits decreases atherosclerotic plaque formation and increases antioxidant status. J. Nutr., 132: 20-26.
PubMedDirect Link - Ryu, S.N., S.Z. Park and C.T. Ho, 1998. High performance liquid chromatographic determination of anthocyanin pigments in some varieties of black rice. J. Food Drug Anal., 6: 729-736.
Direct Link - Saito, K., G.N. Atlin, B. Linquist, K. Phanthaboon, T. Shiraiwa and T. Horie, 2007. Performance of traditional and improved upland rice cultivars under nonfertilized and fertilized conditions in Northern Laos. Crop Sci., 47: 2473-2481.
CrossRef - Shi, W., Y. Yang, S. Chen and M. Xu, 2008. Discovery of a new fragrance allele and the development of functional markers for the breeding of fragrant rice varieties. Mol. Breed., 22: 185-192.
CrossRefDirect Link - Xia, M., W.H. Ling, J. Ma, D.D. Kitts and J. Zawistowski, 2003. Supplementation of diets with the black rice pigment fraction attenuates atherosclerotic plaque formation in apolipoprotein E deficient mice. J. Nutr., 133: 744-751.
PubMed - Xiaodong, X., L. Wenhua, M. Jing, X. Min and H. Mengjun et al., 2006. An anthocyanin-rich extract from black rice enhances atherosclerotic plaque stabilization in apolipoprotein E-deficient mice. J. Nutr., 136: 2220-2225.
CrossRefPubMedDirect Link - Yang, D.S., K.S. Lee, O.Y. Jeong, K.J. Kim and S.J. Kays, 2008. Characterization of volatile aroma compounds in cooked black rice. J. Agric. Food Chem., 56: 235-240.
PubMed - Bradbury, L.M.T., T.L. Fitzgerald, R.J. Henry, Q. Jin and D.L.E. Waters, 2005. The gene for fragrance genotyping in rice. Plant Biotechnol. J., 3: 363-370.
CrossRefDirect Link