
Azarnoosh Jafari
Department of Biology, Faculty of Science, Islamic Azad University, Mashad Campus, Iran
Ali Asghar Maassoumi
Department of Botany, Research Institute of Forests and Rangelands, Tehran, Iran
Mohammad Farsi
Department of Biotechnology, Faculty of Agriculture, Ferdowsi University of Mashhad, Iran
Asian Journal of Plant Sciences
Year: 2008 | Volume: 7 | Issue: 1 | Page No.: 50-59







ABSTRACT
An investigation of karyotype and chromosome numbers was carried out in different populations of Bellevalia Lapeyr. and Muscari Mill. species from Iran. In this research, six species of Bellevalia and six species of Muscari were studied. Different levels of ploidy were found in them. In Bellevalia with x = 4, levels of ploidy were diploid, autopentaploid and hexaploid and in Muscari with x = 9, it was diploid, autotriploid, tetraploid and autopentaploid. In the present research, for first time, the karyotype of B. olivieri was prepared and a population of B. longistyla with autopenploid was observed. The variation of B chromosome number in Bellevalia was considerable.
PDF Abstract XML References Citation
How to cite this article
Azarnoosh Jafari, Ali Asghar Maassoumi and Mohammad Farsi, 2008. Karyological Study on Bellevalia and Muscari (Liliaceae) Species of Iran. Asian Journal of Plant Sciences, 7: 50-59.
DOI: 10.3923/ajps.2008.50.59
URL: https://scialert.net/abstract/?doi=ajps.2008.50.59
DOI: 10.3923/ajps.2008.50.59
URL: https://scialert.net/abstract/?doi=ajps.2008.50.59
INTRODUCTION
Bellevalia Lapeyr. and Muscari Mill. are genera of bulbous plants belonging to Liliaceae, subfamily Lilioideae and tribe Scilleae (Engler, 1887). The both genera have a wide spread distribution. They are present in the whole Mediterranean basin as far as Caucasus, temperate Europe, North of Africa, South west of Asia (Losinskaya, 1935; Feinbrun, 1938-1940; Parsa, 1950; Bentzer, 1972; Garbari, 1973; Davis, 1984; Townsend and Guest, 1985; Feinbrun, 1986; Assadi, 1988; Rechinger, 1990; Wendelbo, 1990).
Bellevalia and Muscari are comprising 18 and nine species in Iran, respectively (Parsa, 1950; Wendelbo, 1967, 1980; Assadi, 1988; Rechinger, 1990). Taxonomically Bellevalia closely is related to Muscari and Hyacinthella. However cytologically, the larger chromosome of Bellevalia make it easily recognizable from the two other ones. The basic chromosome number is x = 4 in Bellevalia (Johnson, 2003) and x = 9 in Muscari (Speta, 1998).
Previous karyological studies on Bellevalia and Muscari showed in Table 1. The length of chromosome of Bellevalia and Muscari species were variable between (6-10, 18-20) and (3-8) micron, respectively (Feinbrun, 1938-1940; Bentzer, 1972).
The aim of this research was to study the karyotype of the mentioned species and to test if the relationship based on morphological characters are in accordance with cytological data. So we prepared karyogram of somatic number of chromosome from B.fominii, B.tabriziana, B. glauca, B.longistyla, B.olivieri, M. comosum, M. caucasicum, M. tenuiflorum, M.longipes, M. armeniacum var. szovitzianum and M. neglectum.
| Table 1: | Previous Karyological studies on studied Bellevalia |
 | |
| Table 2: | Taxa, localities of Bellevalia and Muscari |
 | |
MATERIALS AND METHODS
The materials were collected from the east, center and west of Iran in February until June 2003, 2004 (Table 2). Voucher specimen are deposited in herbarium of Tehran sciences and researches campus For karyotype analysis, a pretreatment at room temperature for three hours was usually applied before fixation of the root tips of six species of Bellevalia and Muscari either in 0.002 M 8-Hydroxyquinoline. After fixation in a cold mixture of ethanol and acetic acid (3:1), the following procedure involved the maceration in 1 N HCl at 60 for 5-8 min, washing in water, cutting off the meristems and squashing them in a drop of 45% acetic acid (Krahulcova, 2003). Chromosomes were described according to Levans terminology (Levan et al., 1964). Karyotypes were compared using total form percentage (Forni-Martin et al., 1994) and calculated the ratio of the longest to the shortest chromosome (Verma, 1980). Symmetry karyotypes were determined using Stebbins two way system (Stebbins, 1971).
RESULTS
The somatic chromosome number and details of karyotype of Bellevalia and Muscari studied species were shown in Table 3 and 4. B. fominii from sect. Nutans (Fig. 1a, 2a) and B. tabrizina from sect. Patens (Fig. 1b, 2b) were diploid (2X = 2n = 8) with karyotype formula
| Table 3: | Somatic chromosome no., karyotype formula and symetoy of Bellevalia and Muscari |
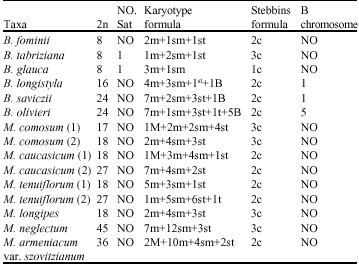 | |
| NO = Not Observed M = Median point, m = Median region, sm = Submedian, st = Subterminal t = Terminal region | |
| Table 4: | Karyotypic details of the species studies of Bellevalua and Muscari |
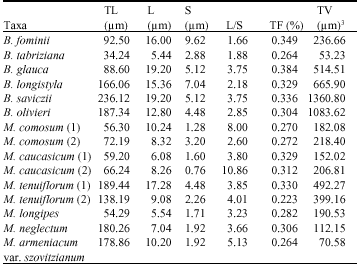 | |
| TL = Total chromatin length, S = Shortest chromosome, L = Longest chromosome, L/S = Longest/Shortest, TF % = Total from percentage, TV = Total Volume | |
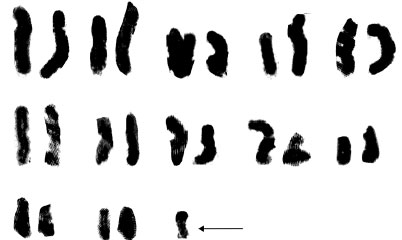
(2m+1sm+1st) and (1m+2sm+1st), respectively. The rest of species belonged to sect. Bellevalia. B. glauca was diploid (2X = 2n = 8) with karyotype formula (3m+1sm) and the satellite was observed above the short arm of first pair chromosome (Fig. 1c, 2c). B. longistyla was autopentaploid (5X = 2n = 20+1B) with sm B-chromosome and karyotype formula (1m+5sm+4st+1B) (Fig. 1d, 2d). B. saviczii was hexaploid (6X = 2n = 24+1B) with sm B-chromosome and karyotype formula (7m+2sm+3st+1B) (Fig. 1e, 2e). B. olivieri was hexaploid (6X = 2n = 24+5B) with five sm, st and t B-chromosome and karyotype formula (7m+1sm+3st+1t+ 5B) (Fig. 1f, 2f).
Also M. comosoum (1) was diploid (2X = 2n = 18) with (1M+2m+2sm+4st) (Fig. 1g, 2g.), but in M. comosum (2) was diploid (2X = 2n = 17) with karyotype formula (2m+4sm+3st) (Fig. 1h, 2h). M. caucasicum (1), (2) were
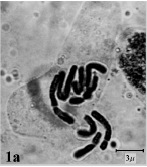 | |
| Fig.1a: | Somatic chromosomes of B. fominii |
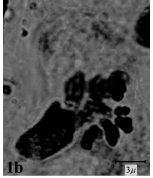 | |
| Fig.1b: | Somatic chromosomes of B. tabriziana |
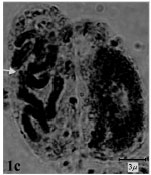 | |
| Fig.1c: | Somatic chromosomes of B. glauca The arrow showing satellite |
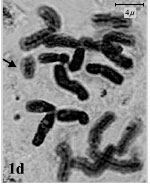 | |
| Fig.1d: | Somatic chromosomes of B. longistyla. The arrow showing B chromosome |
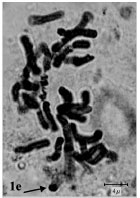 | |
| Fig.1e: | Somatic chromosomes of B. saviczii. The arrow showing B chromosome |
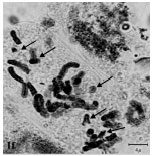 | |
| Fig.1f: | Somatic chromosomes of B. olivieri. The arrows showing B chromosomes |
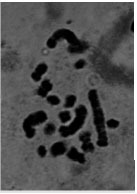 | |
| Fig.1g: | Somatic chromosomes of M. comosum (1) |
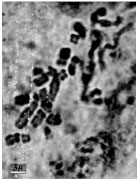 | |
| Fig.1h: | Somatic chromosomes of M. comosum (2) |
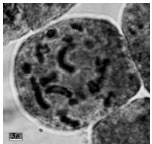 | |
| Fig.1i: | Somatic chromosomes of M. caucasicum (1) |
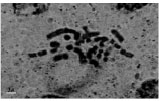 | |
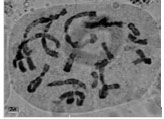
| Fig.1j: | Somatic chromosomes of M. caucasicum (2) |
 | |
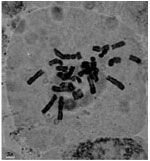
| Fig.1k: | Somatic chromosomes of M. tenuiflorum (1) |
 | |
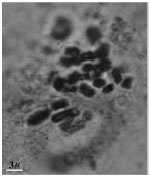
| Fig.1l: | Somatic chromosomes of M. tenuiflorum (2) |
 | |
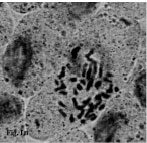
| Fig.1m: | Somatic chromosomes of M. longipes |
 | |
| Fig.1n: | Somatic chromosomes of M. neglectum |
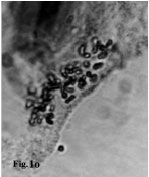 | |
| Fig.1o: | Somatic chromosomes of M. armeniacum var. szovitzianum |
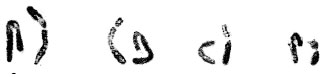 | |
| Fig. 2a: | Karyotype of B. fominii |
 | |
| Fig. 2b: | Karyotype of B. tabriziana |
 | |
| Fig. 2c: | Karyotype of B. glauca |
 | |
| Fig. 2d: | Karyotype of B. longistyla. The arrow showing B chromosome |
 | |
| Fig. 2e: | Karyotype of B. saviczii. The arrow showing B chromosome |
 | |
| Fig. 2f: | Karyotype of B. olivieri |
 | |
| Fig. 2g: | Karyotype of M. comosum (1) |
 | |
| Fig. 2h: | Karyotype of M. comosum (2) |
 | |
| Fig. 2i: | Karyotype of M. caucasicum (1) |
 | |
| Fig. 2j: | Karyotype of M. caucasicum (2) |
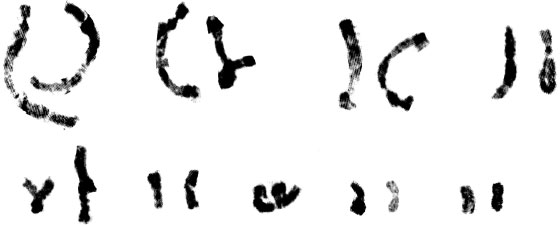 | |
| Fig. 2k: | Karyotype of M. tenuiflorum (1) |
 | |
| Fig. 2l: | Karyotype of M. tenuiflorum (2) |
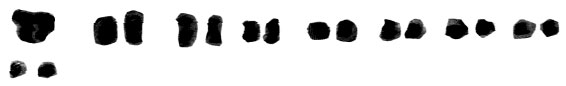 | |
| Fig. 2m: | Karyotype of M. longipes |
 | |
| Fig. 2n: | Karyotype of M. neglectum |
 | |
| Fig. 2o: | Karyotype of M. armeniacum var. szovitzianum |
(2X, 3X = 2n = 18, 27) with karyotype formula (1M+3m+4sm+1st) and (7m+4sm+2st), respectively (Fig. 1i, j, 2i, j). M. tenuiflorum (1), (2) were diploid and autotriploid (2X, 3X = 2n = 18, 27) with karyotype formula (5m+3sm+1st) and (1m+5sm+6st+1t) and some secondary constrictions (Fig. 1k, l, 2k, l). M. longipes was diploid (2x = 2n = 18) with karyotype formula (2m+4sm+3st) (Fig. 1m, 2m). M. neglectum was autopentaploid (5X = 2n = 45) with karyotype formula (7m+12sm+3st) (Fig. 1n, 2n) and M. armeniacum var. szovitzianum was tetraploid (4x = 2n = 36) with karyotype formula (2M+10m+4sm+2st) (Fig. 1o, 2o).
DISCUSSION
The basic chromosome number of Bellevalia is X = 4. This genus often are diploid with 2n = 8, but there is a polyploidy series of 2n = 16, 24, 32. Aneuploidy occurs only at the octaploid level (Ozhatay and Johnson, 1996). They have metacentric, acrocentric and telocentric B-chromosome (Johnson, 2003).
B. fominii was falling to class 2C. This species had the longest chromosome among the studied species which didn`t have any chromosome polymorphism. Also some weak constriction were above the long arms. In Bothmers report, karyotype formulae is the same as the present research. Bothmer reported one chromosome in pair No. 4 have a longer short arm than usual which indicate a pericentric inversion (Bothmer and Wendelbo, 1981). B. tabriziana had the shortest and thickest chromosome among the studied species. This species was falling to class 3C. Its chromosomes were large M and short st while Persson and Wendelbo reported its chromosome have been large M, sm and st (Persson and Wendelbo, 1979). The ploidy level of B. tabriziana in this research paper was similar to Perssons report. B. glauca was falling to class 1C. The satellite was observed above the short arm of first pair chromosome. The chromosome polymorphism was occurred on first pair chromosome. The long arms of first pair were not equal. It is related to pericentric inversion. This species had the biggest and the thickest chromosomes among the studied sect. Bellevalia. B. longistyla was falling to class 2C. The autopentaploid population for this species is recorded for the first time. No obvious chromosome polymorphism was observed. B. saviczii with one B chromosome was falling to class 2C. Its chromosomes were smaller and thinner than the other studied species of sect. Bellevalia in this research. In present paper, allocyclic event and secondary constriction in long arm were observed. Only one sm B-chromosome also, was found while in Bothmers report its number was 1, 3, 7. Getner also reported 1-8 B-chromosome which the most and the least of them were m and M, t (Getner, 2005). The chromosome polymorphism was occurred in pairs No. 2, 3, 4. Getner also confirmed pericentric inversion. He believed the acrocentric was changed to metacentric. In its report aneuploidy, pentasomic (2n-1 = 23), heptasomic (2n+1 = 25) and octasomic (2n+2 = 23) were found in B. saviczii which collected from Shiraz (South of Iran) (Getner, 2005). Zakharieva reported pentaploid population of B. saviczii (Zakhariyeva and Makushenko, 1969). B. olivieri was falling to class 2C. The detail of karyotype is presented for first time. It has five B-chromosomes which were variable as st, sm`, m. Asymmetric karyotype was bimodal. Pericentric inversion and to minor extent translocations seems to be the main background for the chromosome polymorphism (Bothmer and Wendelbo, 1981) while Feinbrun believed allopolyploidy occurred in Bellevalia (Feinbrun, 1938-1940). In regarding to Muscari, M. comosum (1), (2) were diploid and falling to class 3C but first population showed decreasing aneuploidy. Also one of the small chromosomes didn`t have homologous. One of the first pair chromosome was longer than the other long st chromosome. Asymmetric karyotype was trimodal. In Bentzers report, M. comosum was 2n = 18, 27 (Bentzer, 1972). Both of the population of M. caucasicum were diploid and autotriploid and falling to class 2C. Asymmetric karyotype was trimodal. Both of the population of M. tenuiflorum were diploid and autotriploid that falling to class 2C and 3C. Some constrictions were recognized. They had trimodal asymmetric karyotype. In this specimen allocyclic event existed.
| • | M. longipes was diploid and falling to class 3C. |
| • | M. neglectum and M. armeniacum var. szovitzianum were autopentaploid and tetraploid that falling to class 2C |
Most of reports of polyploidy level in Bellevalia showed diploid and tetraploid population. Few reports exists about triploid (Musano and Maggini, 1976) and octaploid populations (Zakhariyeva and Makushenko, 1969; Bothmer and Wendelbo, 1981; Pogosjan and Torosyan, 1983). Variation of ploidy and similarity of morphological characters have been found among the species. In terms of morphological characters, Morphologically, Bellevalia sect. Bellevalia is closer to Muscari subgen. Leopoldia. Also subgen. Botryanthus is more advanced than subgen. Leopoldia. The morphology of chromosomes confirmed taxonomic position. Most of studied Bellevalia species were placed in class 2C of Stebbins system except B. glauca and B. tabriziana.. It is representing more primitive karyotypes.
The karyotypes of both genus are markedly asymmetrical because polymorphism as confirmed by Garbari (1969, 1973), Bentzer and Ellmer (1975), Bentzer and Landstorm (1975), Dalgic (1991), Corsi et al. (1996) and Bareka and Kamari (2001).
Also, data regarding the total chromatin length and size of the longest and shortest chromosomes were shown in Table 4. In Bellevalia studied species, B. saviczii had the highest amount of total chromatin length (236.12 μM) and B. tabrizana had the least (32.24 μM). Also in Muscari, M. tenuiflorum (1) had the highest amount of total chromatin length (189.44) and M. longipes had the least (54.29). Also, M. comosum (2) with 2n = 18 had 72.19 μM total chromatin length while M. caucasicum (2) with 2n = 27 had 66.24 μM.
M. tenuiflorum (1) with 2n = 18 had 189.44 total chromatin length and M. tenuiflorum (2) with 2n = 27 had 138.19. Here M. tenuiflorum (1) with 2n = 18 had 189.44 total chromatin length, M. neglectum with 2n = 36 had 180.26 and M. armeniacum var. szovitzianum with 2n = 45 had 178.86. All of the above cases, are indicating that polyploidy has been accompanied with chromatin loss. Mean while Stebbins class about karyotype was shown in Table 3. Most of studied Bellevalia species were placed in class 2c of Stebbins system except B. glauca and B. tabriziana. It is representing more primitive karyotypes. Most of studied Muscari species were placed in class 2c. Except the both of M. comosum and M. tenuiflorum (2) populations which were placed in 3C because they have trimodal asymmetric karyotype with 3 sizes of chromosomes.
The results were obtained from the present study can be summarized as follows:
| • | Polyploidy variation in Bellevalia was more than Muscari. |
| • | Ploidy increasing didn`t have corresponding to total chromatin length increasing. |
| • | Chromosome morphology of Muscari subgen. Leopoldia is larger and similar to Bellevalia. This subgenus is more primitive than subgen. Botryanthus and closer to Bellevallia (primitive genus). |
| • | In B. longistyla and B. olivieri autopentaploidy and hexaploidy were observed. |
| • | The results of karyological study proved taxonomical position of studied species of sect. Bellevalia. |
| • | Presence of M chromosome in M. armeniacum var. szovitzianum and M. caucasicum (1) probably due to pericentric inversion. |
REFERENCES
- Al-Mudarris, H.E., 1973. Cytological studies on the genus Bellevalia Lapeyr. From Iraq. Bull. Coll. Sci. Univ. Baghdad, 14: 177-195.
Direct Link - Bentzer, B. and Ellmer, 1975. A case of stable chromosomal polymorphism in Leopoldia comosa (Liliaceae). Hereditas, 81: 127-132.
CrossRef - Bentzer, B. and T. Landstrom, 1975. Polymorphism in chromosome of Leopoldia comosa (Liliaceae) revealed by Giemsa staining. Hereditas, 80: 219-239.
CrossRef - Bothmer, R.V. and P. Wendelbo, 1981. Cytological and morphological variation Bellevalia. Nord. J. Bot., 1: 4-11.
CrossRef - D'Amato, F., 1952. New evidence on endopolyploidy in differentiated plant tissue. Caryologia, 4: 121-144.
CrossRefDirect Link - Forni-Martins, E.R., M. Franchi-Tanibata and A. Cardelli-de Lucena, 1994. Karyotypes of species of Sesbania scop. (Fabaceae). Scop. Cytologia, 59: 13-18.
CrossRefDirect Link - Gadella, T.W.J., F. Kliphuis and E.A. Mennega, 1966. Chromosome numbers of some flowering plants of Spain and S. France. Acto. Bot. Nedrl., 15: 484-489.
CrossRefDirect Link - Getner, M.E., 2005. Variable karyotype with B-chromosome in B. saviczii (Liliaceae). Genetica, 124: 223-234.
CrossRefDirect Link - Goldblatt, P., 1974. Chromosome numbers of phanerogams. 5. Ann. Missouri. Bot. Garden. St Louis, 61: 901-904.
Direct Link - Johnson, M.A.T. and P.E. Brandham, 1997. New chromosome numbers in petaloid monocotyledons and in other miscellaneous angiosperms. Kew. Bull., 52: 121-136.
CrossRefDirect Link - Johnson, M.A.T., 2003. Polyploidy and karyotype variation in Turkish Bellevalia (Hyacinthaceae). Bot. J. Linn. Soc., 143: 87-98.
CrossRef - Karlen, T., 1984. Karyotypes and chromosome numbers of five species of Muscari (Liliaceae). Willdenowia, 14: 312-320.
Direct Link - Krahulcova, A., 2003. Chromosome numbers in selected monocotyledons (Czech Republic). Perslia. Praha., 75: 97-113.
Direct Link - Levan, A., K. Fredga and A.A. Sandberg, 1964. Nomenclature for centromeric position on chromosomes. Hereditas, 52: 201-220.
CrossRefDirect Link - Stebbins, G.L., 1971. Chromosomal Evolution in Higher Plants. 1st Edn., Edward Arnold Publisher Ltd., London, UK., Pages: 216.
Direct Link