
A.K.S. Lobato
Instituto de Ciências Agrárias, Universidade Federal Rural da Amazônia, Belém, Brazil
C.F. Oliveira Neto
Instituto de Ciências Agrárias, Universidade Federal Rural da Amazônia, Belém, Brazil
R.C.L. Costa
Instituto de Ciências Agrárias, Universidade Federal Rural da Amazônia, Belém, Brazil
B.G. Santos Filho
Instituto de Ciências Agrárias, Universidade Federal Rural da Amazônia, Belém, Brazil
F.J.R. Cruz
Instituto de Ciências Agrárias, Universidade Federal Rural da Amazônia, Belém, Brazil
H.D. Laughinghouse IV
Department of Biology, John Carroll University, University Heights, OH, USA
Asian Journal of Plant Sciences
Year: 2008 | Volume: 7 | Issue: 1 | Page No.: 44-49







ABSTRACT
The aim of this experiment was to investigate the responses caused by progressive water stress and the necessary time for have biochemical and physiological changes of Vigna unguiculata (L.) Walp. cv: manteguinha during the vegetative phase. Leaf Relative Water Content (LRWC), Nitrate Reductase Activity (NRA), free proline, total soluble carbohydrates, free amino acids and total soluble proteins were quantified. The plants under the control treatment maintained stable variables; however, those under water stress suffered an increase of 97.3% in proline due to osmotic adjustment, increase of 78.4% in free amino acids, caused by the break down of proteins by protease enzymes, increase in total carbohydrates by 94.2% due to a decrease in the photosynthetic capacity and low synthesis of sucrose used for exporting this solute. There was a decrease in 25.7% of the leaf relative water content in virtue of low water availability in the soil, reduction of 43.8% in Nitrate Reductase Activity (NRA) due to the regulation of this enzyme being induced by its substratum and 47.3% of total proteins due to the paralyzation of protein and proteolytic protein degradation, being maximized the variations in the parameters evaluated according to the increase in the period the plants were exposed to water stress. Physiological changes can be carried out in only a few days. However, biochemical responses should be evaluated starting at 4 days under water restriction.
PDF Abstract XML References Citation
How to cite this article
A.K.S. Lobato, C.F. Oliveira Neto, R.C.L. Costa, B.G. Santos Filho, F.J.R. Cruz and H.D. Laughinghouse IV, 2008. Biochemical and Physiological Behavior of Vigna unguiculata (L.) Walp. Under Water Stress During the Vegetative Phase. Asian Journal of Plant Sciences, 7: 44-49.
DOI: 10.3923/ajps.2008.44.49
URL: https://scialert.net/abstract/?doi=ajps.2008.44.49
DOI: 10.3923/ajps.2008.44.49
URL: https://scialert.net/abstract/?doi=ajps.2008.44.49
INTRODUCTION
Water availability is considered the climatic factor with greatest effect on agricultural productivity, being responsible to govern species distribution in the different climate zones around the globe (Rockström and Falkenmark, 2000). The effects of drought vary and depend on the intensity, development stage and duration of the hydric stress, as well as, the adaptive strategies that the species possess to tolerate this abnormal condition (Kramer and Boyer, 1995).
During the vegetative phase, water deficit causes leaf and plant growth reduction, besides altering the process of nutrient absorption due to the low water availability in the environment and low photosynthetic activity, since with water stress an increase in stomatal resistance occurs, decreasing the capacity of gaseous exchange between the environment and the plant (Kerbauy, 2004).
The water deficit influences various biochemical and physiological processes (Sircelj et al., 2005), where proline and carbohydrates constitute some of the organic solutes, which accumulate in the plant cells for adjusting plant osmosis (Taiz and Zeiger, 1998; Zhang et al., 1999). Many researchers have reported alterations in the functioning and speed of enzymatic activity, like nitrate reductase, amino acid synthesis (Andrews et al., 2004) and decrease in protein levels (Zhu and Xiong, 2002), as metabolical responses to water restrictions (Pimentel, 2004).
Cowpea (Vigna unguiculata (L.) Walp.) is an edible dicotiledoneous legume with the capacity to store large quantities of protein in its grain, having an excellent capacity at fixing nitrogen, as well as not needing very fertilized soil (Lobato et al., 2006; Peksen and Artik, 2004).
Studies indicate that cowpea tolerates low water availability and high temperatures, besides revealing that the species supports up tp 10 days under water defficiency, simulated in a greenhouse (Costa, 1999; Silveira et al., 2001, 2003); however, the consequences on the physiological and biochemical metabolisms involved in conditions of light, moderate and severe stress are not well-known for manteguinha cultivars.
The aim of the experiment was to investigate the responses caused by progressive water stress and the necessary time for have changes on the LRWC, NRA, proline, total soluble carbohydrates, free amino acids and total soluble proteins of plants of Vigna unguiculata (L.) Walp. cv: manteguinha during the vegetative phase.
MATERIALS AND METHODS
The experiment was conducted in a greenhouse, located at the Instituto de Ciências Agrária (ICA) of the Universidade Federal Rural da Amazônia (UFRA), city of Belém, state of Pará, Brazil (0127`S and 4826`W), during the months of September and October of 2006, under natural conditions: day/night (minimum/maximum air temperature and relative humidity were: 22.4/37.6C and 68/79%, respectively, verified during the experiment), where the average photoperiod was 12 h and the maximum active photosynthetic radiation, 525 μmol-2 sec–1 (at 12:00 h).
The plants were grown in 6 L pots filled with black potting soil and aviary manure, mixed at 3:1 proportion. Vigna unguiculata seeds collected in the 2006 season were stored until the experiment and used.
The experimental design was randomized, with 2 water conditions (stress and control) and 4 evaluation points (0, 3, 6 and 9 days), with 8 repetitions and 64 experimental units, where each repetition was composed by one plant.
Three seeds were placed into each pot and after 7 days, the plants were thinned to one per pot only. The plants remained in a greenhouse for 35 days, watered daily and received macro and micronutrients every 5 days, using the nutritive solution by Hoagland and Arnon (1950). Starting the 35th day after the implementation of the experiment, the plants from the treatment under stress were submitted to a period of 9 days without irrigation, until the 44th day, simulating water deficit (stress).
Physiological analyses: The plants were taken to the Laboratory of Advances Plant Physiology (UFRA) to perform the analyses. The nitrate reductase activity was performed with disks of fresh leaves measuring 0.5 cm2 in area and the spectrophotometer readings were 540 nm (Hageman and Hucklesby, 1971). The determination of the Leaf Relative Water Content (LRWC) was performed with 10 mm disks in diameter, as well as RWC was calculated as:
LRWC = [(FW-DW)/(TW-DW)]x100
Where:
FW=The fresh weight
TW=Turgid weight measured after 24 h of saturation in de-ionized water at 4C in the dark and DW is the dry weight determined after 48 h in an oven at 80C (Slavick, 1979)
Biochemical analyses: The fresh leaves were oven dried at 65C for 48 h, until obtaining dry matter for quantification in the spectrophotometer. The free proline was determined at 520 nm (Bates et al., 1973), free amino acids at 570 nm (Peoples et al., 1989), total soluble carbohydrates at 490 nm (Dubois et al., 1956) and total soluble proteins at 595 nm (Bradford, 1976).
Statistical analyses: The data was submitted to ANOVA and when there was a significant difference, the Tukey test at 5% significance was applied, as well as was calculated standard error for each point, where the statistical analyses were performed with SAS (1996).
RESULTS
Leaf relative water content and nitrate reductase activity: The LRWC suffered a significant reduction of 25.7%, according to the variance analysis, in which the treatment under stress decreased from 89.7 to 66.7% at 0 and 9 days of stress, respectively (Fig. 1A). This reveals that an
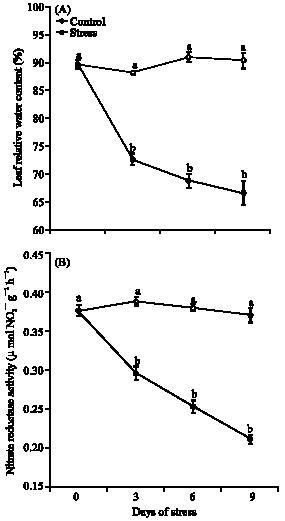 | |
| Fig. 1: | Relative water content (A) and nitrate reductase activity (B) in plants of V. unguiculata under 0, 3, 6 and 9 days of hydric stress. Averages followed by the same letter(s) do not differ among themselves by the Tukey test at 5% of probability and the bars represent the mean standard error |
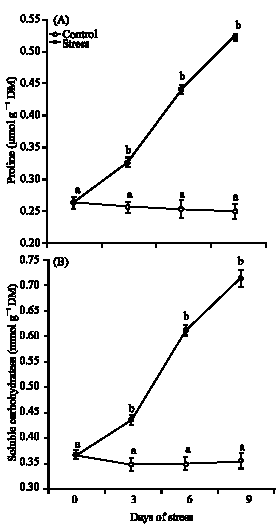 | |
| Fig. 2: | Tenors of free proline (A) and total soluble carbohydrates (B) in plants of V. unguiculata under 0, 3, 6 and 9 days of hydric stress. Averages followed by the same letter do not differ among themselves by the Tukey test at 5% of probability and the bars represent the mean standard error |
accentuated drop occurred in this variable during the period between 0 and 3 days, contributing with 19% of the reduction total. However, the control treatment remained stable and above the stress, varying between 88 and 91%.
The NRA decreased significantly, 43.8% (Fig. 1B), when the period under water restriction increased (9 days). It has been observed that there is a relation between the relative water content and nitrate reductase, because the larger variation occurred during the period of 0 to 3 days, with 21.4% of the total reduction. As well as similar to what was observed with the LRWC. The control kept the levels of nitrate reductase stable. Our results are in agreement with Marur et al. (2000), who worked with Grossypium hirsutum and Chandrasekar et al. (2000), studying Triticum spp. on the influence of abiotic stresses in the reduction of enzyme activity.
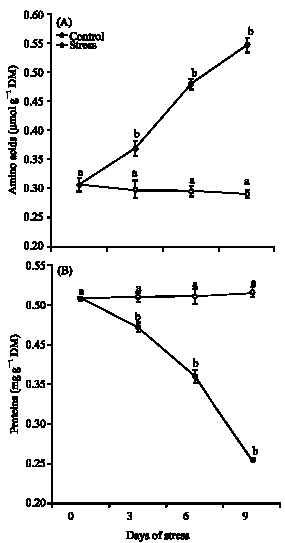 | |
| Fig. 3: | Tenors of free amino acids (A) and total soluble proteins (B) in plants of V. unguiculata under 0, 3, 6 and 9 days of hydric stress. Averages followed by the same letter do not differ among themselves by the Tukey test at 5% of probability and the bars represent the mean standard error |
Proline and carbohydrates: In the plants submitted to 9 days of water stress, a significant increase of 97.3% in free proline was observed (Fig. 2A). It was seen that in the period between 3 and 6 days a larger variation, 43% increase, occurred in the proline while under water stress. Similar results were observed by Costa (1999) investigating Vigna unguiculata.
Carbohydrate levels suffered a significant increase of 94.2% in the plants submitted to 9 days of water stress (Fig. 2B), in which 48% of the increase was observed in the period of 3 to 6 days under water restriction; however the plant carbohydrate levels under the control treatment remained stable.
Amino acids and proteins: A significant elevation of 78.4% in the levels of free amino acids occurred under
9 days of water restriction, when the major period increased from 3 to 6 days with 36.3%. However, the control treatment showed stable levels of amino acids (Fig. 3A). These results corroborate with the studies by Silveira et al. (2001) with Vigna unguiculata under saline stress and Pimentel (1999) on the behavior of Zea mays under water stress.
Significant reduction, 47.3%, in the levels of total soluble proteins under 9 days of water stress, as well as the period between 6 and 9 days showed a major drop and contributed with 24.3% of the total reduction. The control maintained little variation in the protein levels (Fig. 3B).
DISCUSSION
The drop in the leaf relative water content is a result of low water availability in the soil, where the osmosis in the environment was progressively becoming negative, causing many biochemical and physiological alterations which aimed at decreasing the plant water loss to the environment during transpiration, to maintain the metabolic function and to adjust the species osmotically (Kerbauy, 2004).
The nitrate reductase enzyme activity decreases due to a drop in nitrate absorption carried out by plants under water stress; hence the inorganic nitrogen is assimilated by the root system using water as a vehicle (Larsson, 1992). Nitrate reductase is considered an excellent physiological indicator, since it is the first enzyme in the nitrogen metabolism, as well as its activity being regulated by NO3– and glutamine, which is the final product of the metabolic nitrogen pathway (Foyer et al., 1998; Ferrario-Méry et al., 1998). These results reveal that the plants under water stress assimilate small quantities of nitrogen (Crawford, 1995). Under normal conditions the nitrate is used to form proteins, nucleic acids and other cellular components, this being the biggest source of nitrogen for vascular plants (Ezzine and Ghorbel, 2006).
The increase of free proline in this experiment is attributed to the osmotic adjustment (Verslues et al., 2006), in which V. unguiculata, under water stress, has low hydric potential and in reply elevated the levels of proline aiming at tolerating the abnormal situation to which it is submitted (Vendruscolo et al., 2007). Since proline is an organic solute, it accumulates in the cytoplasm, protecting cellular structures and macromolecules from denaturation (Vanrensburg et al., 1993), aside from being a source of energy (Serrano and Gaxiola, 1994). On the other hand, the plants in the control treatment maintained the levels of proline low and stable, because free proline is easily converted into glutamic acid and other components, under the normal irrigation conditions (Sarker et al., 1999).
The total soluble carbohydrates were progressively accumulated in the plant leaves during water stress, due, initially, to the hydric restriction. This provoked a reduction in the quantity of starch by the amylase enzyme activity and increase in the stress level (Chaves Filho and Stacciarini-Seraphin, 2001), probably occurring due to the accumulation of other soluble carbohydrates, principally fructose, triose and glucose, besides sacarose in lower quantity compared to the other sugars, being responsible of elevating this variable even more (Lawlor and Cornic, 2002).
The decrease observed in soluble proteins occurred primarily due to the paralyzation of protein biosynthesis (Kramer and Boyer, 1995) and as the period under water restrictions increases, a proteolytic degradation of proteins occurs (Roy-Macauley et al., 1992) decreasing, even further, the protein levels. It has to be taken into consideration that between the proteins present in plants, rubisco (ribulose 1,5 bisphosphate carboxilase-oxigenase) is the most important, because it composes more than 50% of the leaf cellular protein and constitutes the biggest nitrogen reserve in plant tissues, aside from being fundamental in the photosynthesis process (Lawlor, 2002). Studies conducted by Flexas et al. (2006) indicate that the activity regulation of this protein is correlated with the stomatal conductance, being observed an accentuated drop plant activity under water stress caused by higher stomatal resistance.
An increase in free amino acids was observed, where the increase was attributed to the damages caused by the water deficit, which initially promoted the increase of this metabolic provoked by protein degradation by proteases. With the increase in water stress severity, a degradation of the enzymatic systems occurred, where it was necessary to synthesize the amino acids again, which made the amino acid concentration in the leaves even more elevated with the objective of osmotically adjusting the plant under hydric deficit conditions (Yordanov et al., 2000; Hoekstra et al., 2001).
This study revealed that experiments aiming to obtain physiological changes in V. unguiculata can be carried out in only a few days. However, the biochemical responses should be evaluated starting at 4 days under water restriction.
REFERENCES
- Bates, L.S., R.P. Waldren and I.D. Teare, 1973. Rapid determination of free proline for water-stress studies. Plant Soil, 39: 205-207.
CrossRefDirect Link - Bradford, M.M., 1976. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem., 72: 248-254.
CrossRefPubMedDirect Link - Chandrasekar, V., R.K. Sairam and G.C. Srivastava, 2000. Physiological and biochemical responses of hexaploid and tetraploid wheat to drought stress. J. Agron. Crop Sci., 185: 219-227.
CrossRefDirect Link - Chaves Filho, J.T. and E. Stacciarini-Seraphin, 2001. [Changes in osmotic potential and soluble carbohydrates levels in Solanum lycocarpum St.-Hil. in response to water stress]. Braz. J. Bot., 24: 199-204, (In Portuguese).
CrossRefDirect Link - Crawford, N.M., 1995. Nitrate: Nutrient and signal for plant growth. Plant Cell, 7: 859-868.
PubMedDirect Link - DuBois, M., K.A. Gilles, J.K. Hamilton, P.A. Rebers and F. Smith, 1956. Colorimetric method for determination of sugars and related substances. Anal. Chem., 28: 350-356.
CrossRefDirect Link - Ezzine, M. and M.H. Ghorbel, 2006. Physiological and biochemical responses resulting from nitrite accumulation in tomato (Lycopersicon esculentum Mill. Cv. Ibiza F1). J. Plant Phys., 163: 1032-1039.
CrossRefPubMedDirect Link - Ferrario-Mery, S., M.H. Valadier and C.H. Foyer, 1998. Overexpression of nitrate reductase in tobacco delays drought-induced decreases in nitrate reductase activity and mRNA. Plant Physiol., 117: 293-302.
PubMed - Flexas, J., M. Ribas-Carbo, J. Bota, J. Galmes, M. Henkle, S. Martinez-Canellas and H. Medrano, 2006. Decreased Rubisco activity during water stress is not induced by decreased relative water content but related to conditions of low stomatal conductance and chloroplast CO2 concentration. New Phytol., 172: 73-82.
CrossRefPubMedDirect Link - Foyer, C.H., M.H. Valadier, A. Migge and T.W. Becker, 1998. Drought-induced effects on nitrate reductase activity and mRNA and on the coordination of nitrogen and carbon metabolism in maize leaves. Plant Phys., 117: 283-292.
CrossRefDirect Link - Hageman, R.H. and D.P. Hucklesby, 1971. Nitrate reductase from higher plants. Methods Enzimol., 23: 491-503.
CrossRefDirect Link - Hoekstra, F.A., E.A. Golovina and J. Buitink, 2001. Mechanisms of plant desiccation tolerance. Trends Plant Sci., 6: 431-438.
CrossRefPubMedDirect Link - Larsson, M., 1992. Translocation of nitrogen in osmotically stressed wheat seedling. Plant Cell Environ., 15: 447-453.
CrossRef - Lawlor, D.W., 2002. Carbon and nitrogen assimilation in relation to yield: Mechanisms are the key to understanding production systems. J. Exp. Bot., 53: 773-787.
CrossRefDirect Link - Lawlor, D.W. and G. Cornic, 2002. Photosynthetic carbon assimilation and associated metabolism in relation to water deficits in higher plants. Plant Cell Environ., 25: 275-294.
CrossRefDirect Link - Marur, C.J., P. Mazzafera and A.C. Magalhaes, 2000. Nitrate reductase activity in cotton plants under water deficit and after turgescence recovery. Sci. Agric., 57: 277-281.
CrossRefDirect Link - Peksen, E. and C. Artik, 2004. Comparison of some cowpea (Vigna unguiculata L. Walp) genotypes from Turkey for seed yield and yield related characters. J. Agron., 3: 137-140.
CrossRefDirect Link - Pimentel, C., 1999. Water relations in two hybrids of cornunder two cycles of water stress. Pesq. Agropec. Bras., 34: 2021-2027.
CrossRef - Rockstrom, J. and M. Falkenmark, 2000. Semiarid crop production from a hydrological perspective: Gap between potential and actual yields. Crit. Rev. Plant Sci., 19: 319-346.
CrossRefDirect Link - Roy-Macauley, H., Y. Zuily-Fodil, M. Kidric, A.T. Pham Thi and J.V. de Silva, 1992. Effect of drought stress on proteolytic activities in Phaseolus and Vigna leaves from sensitive and resistant plants. Physiol. Plant., 85: 90-96.
CrossRefDirect Link - Sarker, A.M., M.S. Rahman and N.K. Paul, 1999. Effect of soil moisture on relative leaf water content, chlorophyll, proline and sugar accumulation in wheat. J. Agron. Crop Sci., 183: 225-229.
CrossRefDirect Link - Serrano, R. and R. Gaxiola, 1994. Microbial models and salt stress tolerance in plants. Crit. Rev. Plant Sci., 13: 121-138.
Direct Link - Silveira, J.A.G., A.R.B. Melo, R.A. Viégas and J.T.A. Oliveira, 2001. Salinity-induced effects on nitrogen assimilation related to growth in cowpea plants. Environ. Exp. Bot., 46: 171-179.
CrossRefDirect Link - Silveira, J.A.G., R.C.L. Costa, R.A. Viegas, J.T.A. Oliveira and M.V.B. Figueiredo, 2003. N-Compound accumulation and carbohydae shortage on N2 fixation in drought-stressed and rewatered cowpea plants. Spanish J. Agric. Res., 1: 65-75.
Direct Link - Van Rensburg, L., G.H.J. Kruger and H. Kruger, 1993. Proline accumulation the drought-tolerance selection: Its relationship to membrane integrity and chloroplast ultra structure in Nicotiana tabacum L. J. Plant Physiol., 141: 188-194.
CrossRefDirect Link - Verslues, P.E., M. Agarwal, S. Katiyar-Agarwal, J. Zhu and J.K. Zhu, 2006. Methods and concepts in quantifying resistance to drought, salt and freezing, abiotic stresses that affect plant water status. Plant J., 45: 523-539.
CrossRefPubMedDirect Link - Yordanov, I., V. Velikova and T. Tsonev, 2000. Plant response to drought, acclimation and stress tolerance. Photosynthetica, 38: 171-186.
CrossRefDirect Link - Zhang, J., H.T. Nguyen and A. Blum, 1999. Genetic analysis of osmotic adjustment in crop plants. J. Exp. Bot., 50: 291-302.
CrossRefDirect Link - Xiong, L. and J.K. Zhu, 2002. Molecular and genetic aspects of plant responses to osmotic stress. Plant Cell Environ., 25: 131-139.
CrossRefPubMedDirect Link - Andrews, M., P.J. Lea, J.A. Raven and K. Lindsey, 2004. Can genetic manipulation of plant nitrogen assimilation enzymes result in increased crop yield and greater N-use efficiency? An assessment. Ann. Applied Biol., 145: 25-40.
CrossRef