
L. Naraghi
Department of Plant Disease Research, Iranian Research Institute of Plant Protection,P.O. Box 1452, Tehran 19395, Iran
A. Heydari
Department of Plant Disease Research, Iranian Research Institute of Plant Protection,P.O. Box 1452, Tehran 19395, Iran
F. Azaddisfani
Agricultural Research and Natural Resource Center of Razavi Khorasan, Mashhad, Iran
Asian Journal of Plant Sciences
Year: 2008 | Volume: 7 | Issue: 4 | Page No.: 389-393







ABSTRACT
In this study, Talaromyces flavus, a fungal antagonist, was isolated from soil samples that were collected from cotton fields in Golestan province of Iran. Antagonistic effects of non-volatile extracts of T. flavus isolates on Verticillium dahliae growth were investigated separately in laboratory experiments. Different isolates of T. flavus caused variable percents of growth inhibition of V. dahliae. Among isolates, ten of them that caused more growth inhibition of V. dahliae than other isolates, were selected for greenhouse experiments. In this step, first inoculum of microsclerotia of V. dahliae and seed treatments affected by non-volatile extracts and ascospores suspension of ten isolates of T. flavus were prepared. For comparison of infection index in seed treatment, greenhouse experiment was performed as a completely randomized design with forty-three treatments and three replications. Results of disease evaluation showed that among seed treatments affected by ascospores suspension of T. flavus isolates, seed treatments of isolates TF-2, Tf-12 and TF-17 caused maximum decrease in Verticillium wilt index. However, among seed treatments affected by non-volatile extracts of T. flavus isolates, seed treatments of isolates TF-1, TF-17 and TF-13 resulted in maximum decrease in disease index.
PDF Abstract XML References Citation
How to cite this article
L. Naraghi, A. Heydari and F. Azaddisfani, 2008. Study on Antagonistic Effects of Non-Volatile Extracts of Talaromyces flavus on Cotton Verticillium Wilt Disease. Asian Journal of Plant Sciences, 7: 389-393.
DOI: 10.3923/ajps.2008.389.393
URL: https://scialert.net/abstract/?doi=ajps.2008.389.393
DOI: 10.3923/ajps.2008.389.393
URL: https://scialert.net/abstract/?doi=ajps.2008.389.393
INTRODUCTION
Cotton is an important cash crop around the world including Iran. Cotton is cultivated in about 20 provinces of Iran (Heydari et al., 2005; Naraghi et al., 2007). Harmful pests (insects, weeds and pathogens) are among the most important yield reducing agents in cotton fields (Niederhauser and Niederhauser, 2000; Mi et al., 1998; Heydari et al., 2005; Heydari and Misaghi, 1998; Naraghi et al., 2007; Abd-Elsalam et al., 2007).
Verticillium wilt is considered the most important disease of cotton in Iran (Heydari et al., 2007; Naraghi et al., 2007). For controlling Verticillium wilt of cotton, cultural practices and the use of resistant varieties are the most common strategies which are not either available or effective. Biological control using fungal and bacterial antagonists in recent years, have been applied to control cotton diseases (Heydari et al., 1997, 2007; Heydari and Misaghi, 1998, 2003; Spink and Rowe, 1989; Aziz et al., 1997).
Talaromyces flavus (Klocker) Stulk and Samson is a fungal antagonist that has been used in biological control of Verticillium wilt of several plants (Marois et al., 1984; Tjamos and Fravel, 1995; Naraghi et al., 2004). Marois et al. (1982) showed that T. flavus decreased the incidence of Verticillium wilt and increased yield of egg-plant in England. Fahima and Henis (1997) showed that T. flavus decreased Verticillium wilt disease on egg-plant at the rate of 77%. The ability of this fungus for occupying rhizosphere of potato, cotton, egg-plant and decreasing germination of microsclerotia of Verticillium dahliae have been reported by Marois et al. (1984).
This fungus (sexual form of Penicillin dangerdii) also was reported as parasite of sclerotia of Rhizoctonia solani and Sclerotinia sclerotiorum (McLaren et al., 1982). Tjamos and Fravel (1997) showed that T. flavus decreased Verticillium wilt on tomato plants. T. flavus and Aspergillus terreus have also been reported to be the inhibitory agents of Verticillium wilt of olive trees in Greece (Tjamos, 1991). Forty percent of non-volatile extracts (Talaron) of T. flavus belongs to Glucose oxidase enzyme and the production of Hydrogen peroxide by this enzyme gives antibacterial and antifungal characteristics to this fungus (Kim and Fravel, 1990).
The objective of this study was to compare two methods of cotton seed treatments with T. flavus ascospore suspension in biological control of Verticillium wilt disease on cotton.
MATERIALS AND METHODS
The following experiments were conducted during year 2005.
Isolation of Talaromyces flavus from soil: Since T. flavus is a thermophilic fungus (Guarro et al., 1999), therefore cotton fields in Karkandeh Research Station in Golestan province of Iran was selected for soil collection. Selective medium (TF medium) was adopted for isolation of these isolates from soil. This medium contained 1 L distilled water, 39 g Potato Dextrose Agar (PDA), 2.0 mL of a 50% solution of lactic acid, 100 mg streptomycin sulfate, 50 mg chlorotetracycline HCl, 50 mg chloramphenicol, 4 mg pimaricin (2.5% sterile water suspension), 30 mg nystatine (Mycostatin, 4960 units mg-1) and 0.5 g oxgall (Bile, bovine). Lactic acid and the antimicrobial agents were added as aqueous solutions to autoclaved PDA at about 50°C. The medium was poured in to petri dishes (18 mL per dish). One milliliter of aliquots was removed from 10-2 to 10-3 dilutions (soil in water) during agitation with a magnetic stirrer and was spread on the TF medium (five plates per replication). Plates were incubated in the dark at 30°C for 7-8 days. T. flavus isolates were detected and identified on the TF medium based on their colony morphology 10 days after incubation. Using the above- described procedure, propagules of T. flavus were isolated from soil (Marois et al., 1984).
Investigation of the antagonistic effects of non-volatile extracts of T. flavus on V. dahliae in laboratory experiments: For preparation of non-volatile extracts, 20 different isolates of T. flavus were cultured on C´Zapeck Dex Broth medium separately and placed on shaker in 50 rpm for ten days. In next step, the culture media were filtered through filter paper with pores in 0.45 μ diameter (Eziashi, 2006). The culture filtrates of fungal isolates were then mixed with PDA medium in 20% concentration separately. Culture filtrates mixed with PDA medium were transferred in to Petri dishes. V. dahliae which was previously isolated from cotton field soil (Naraghi et al., 2007) was cultured on medium surface in every Petri dish when medium temperature attained to 50°C. After one week, diameter of V. dahliae colonies were recorded and growth inhibition percents were evaluated as follows:
 |
Preparation of V. dahliae inoculum for greenhouse experiment: In this experiment, one isolate was used (Naraghi et al., 2007). Preparation of V. dahliae microsclerotia suspension of every strain was carried out separately according to the procedure described by Tjamos and Fravel (1995).
Pot soil inoculation was carried out as follows:
Two hundreds microsclerotia were used for each gram of pot soil. One milliliter of suspension containing microsclerotia was poured in to a 2 cm diameter petri plate and the number of microsclerotia was determined using stereo-dissecting microscope.
Preparation of seed treatment affected by T. flavus for pot soil: In this step, 10 isolate of T. flavus that showed more inhibitory effects on V. dahliae growth were selected for preparation of seed treatments. Cotton seed treatments were divided to two groups. First group was affected by ascospores suspension of T. flavus (2x102 ascospore mL-1) and other group was affected by non-volatile extract of T. flavus. Preparation of non-volatile extracts was described in above section.
For preparation of ascospores suspension of T. flavus, this fungus was cultured on Potato Dextrose Agar (PDA) medium and incubated in 30°C for three weeks. In next step, 10 mL SDW was spread on medium surface containing ascospores and ascospores number of obtained suspension were determined in one milliliter (Marois et al., 1984). This step was carried out for every T. flavus isolate separately.
Evaluation of the antagonistic effects of T. flavus as seed treatment on pathogenicity of V. dahliae in the greenhouse: A completely randomized design with 22 treatments and 3 replications was used. Each replication consisted of a 9 cm diameter and 20 cm height plastic pot containing 3 kg sterile soil of cotton field in Golestan province and five seeds of Varamin cotton cultivar. Treatments consisted of:
| • | Soil and seed without inoculum (negative control) |
| • | Soil with V. dahliae inoculum and seed without inoculum (positive control) |
From 3 to 12-soil with V. dahliae inoculum and seeds treated with non-volatile extracts of 10 T. flavus isolates.
From 13 to 22-soil with V. dahliae inoculum and seeds treated with ascospore suspension of 10 T. flavus isolates.
Leaf Infection Index (Iakutkin and Popov, 1979) was determined 8 weeks after sowing. Analysis of variance and mean comparison were performed using MSTATC statistical software.
RESULTS
Isolation of Talaromyces flavus from soil: As Table 1 shows twenty T. flavus isolates were obtained from cotton fields soil in different regions of Golestan province.
Investigation of the antagonistic effects of non-volatile extracts of T. flavus on V. dahliae in laboratory experiments: As Table 2 shows variable inhibitory percents on V. dahliae growth were induced by different T. flavus isolates. Maximum inhibitory percent was mediated by isolate TF-1 and minimum inhibitory percent belonged to isolate TF-16.
Selected isolates for greenhouse experiments included: TF-1, TF-17, TF-13, TF-2, TF-12, TF-3, TF-18, TF-20, TF-6 and TF-4.
Evaluation of the antagonistic effects of T. flavus as seed treatment on pathogenicity of V. dahliae in the greenhouse: As tables show seed treatments affected by ascospore suspension or non-volatile extracts of T. flavus isolates were placed in 9 statistical groups (Table 3, 4). Minimum infection index mean was 0 that belonged to treatments affected by non-volatile extracts of isolates
| Table 1: | Code of T. flavus isolates obtained from different regions of Golestan province |
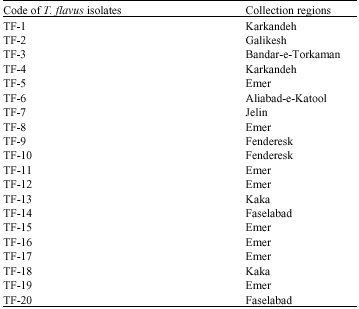 | |
of isolates TF-2, TF-12 and Tf-17 (Table 4). However, there was maximum infection index mean (26.9) in treatment affected by non-volatile extracts of isolate TF-4 (Table 3).
| Table 2: | Inhibitory percents on V. dahliae growth mediated by non-volatile extracts of T. flavus isolates |
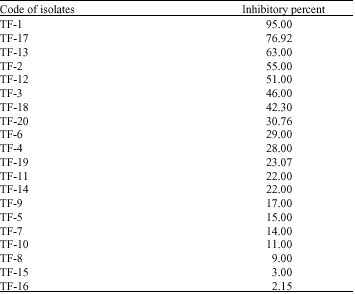 | |
| Table 3: | The effect of seed treatment affected by non-volatile extracts of T. flavus isolates on Verticillium wilt infection index on cotton plants |
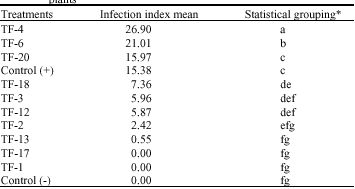 | |
| *Treatments marked by the same letter(s) are not significantly different (p>0.01) | |
| Table 4: | :The effect of seed treatment affected by ascospore suspension of T. flavus isolates on Verticillium wilt infection index on cotton plants |
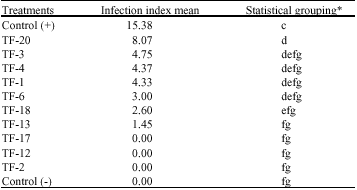 | |
| *Treatments marked by the same letter(s) are not significantly different (p>0.01) | |
DISCUSSION
The overall results of this study show that it may be possible to manage cotton Verticillium wilt disease efficiently by seed treatment with non-volatile extracts or ascospore suspension of T. flavus. The inhibitory effect of metabolites of several bacterial and fungal microorganisms on growth and activity of important fungal pathogens have been shown in previous studies. For example, the efficiency of non-volatile metabolites of some bacteria and fungi such as Aspergillus flavus, A. ochraceus, Penicillium aurantiogriseum, Bacillus subtilis and Trichoderma harzianum for controlling bean antracnose disease have been reported (Adebanjo and Bankole, 2004).
In an other study, the effect of these metabolites on root rot disease of lettuce (Lactuca sativa) caused by Sclerotinia minor was shown in laboratory and greenhouse experiments. This study confirmed that the inhibitory effect of variable components of metabolites on pathogenic agents growth was different (EL- Tarabily et al., 2000). According to conclusion of this study, among these components, chitinase enzyme produced by T. harzianum (Menendez and Godeas, 1998) and T. flavus (Madi et al., 1997) were effective for controlling soybean stem white rot disease caused by S. sclerotiorum and bean stem rot caused by S. rolfsii respectively. Other examples, such as B-1,3 gluconase secreted by Burkholderia cepacia (Fridlender et al., 1993) and glucose oxidase produced by T. flavus (Murrary et al., 1997) have been effective for decreasing seedling damping-off and Verticillium wilt diseases respectively. According to the above studies, for controlling different pathogens, used effective components are variable.
In other study, it has been shown that peroxide hydrogen prevented formation of microsclerotia of V. dahliae and S. rolfsii (Kim and Fravel, 1990). Since, glucose oxidase enzyme produced by T. flavus acts in glucose presence and this activity is resulted in peroxide hydrogen production therefore, glucose addition to non-volatile metabolite of T. flavus caused increase in inhibitory effect on microsclerotia formation (Murrary et al., 1997).
The differences between results of laboratory experiments of this study and the previous ones could be due to metabolite concentration. In this regard, it was reported that there was a minimum concentration for every effective component. For example, minimum concentration of effective component of non-volatile extracts of Fusarium oxysporum (Cyclosporin) used for growth inhibition of S. sclerotiorum was 0.1 μg mL-1 (Rodriguez et al., 2006).
In an other part of this study, using non-volatile extracts and ascospore suspension of T. flavus decreased Verticillium wilt infection index in greenhouse conditions. Results of present study in this part agree with those of some previous studies (Hashem and Ali, 2004; Chitarra et al., 2004). The differences between the results of this study and those of the previous ones could be due to method of seed treatment with non-volatile extracts of T. flavus in intervals before sowing (Hashem and Ali, 2004).
Based on the overall results of this study, it is concluded that the use of antagonistic fungi including T. flavus and its metabolites and ascospore suspension could be an effective method for controlling Verticillium wilt disease of cotton and may be considered a major component of Integrated Pest Management (IPM) strategies for managing this disease which is a very important and destructive disease of cotton in all cotton growing area in the world.
REFERENCES
- Abd-Elsalam, K.A., M.R. Omar, A.R. El-Samavaty and A.A. Aly, 2007. Response of commercial cotton cultivars to Fusarium solani. Plant Pathol. J., 23: 62-69.
Direct Link - Adebanjo, A. and S.A. Bankole, 2004. Evaluation of some fungi and bacteria for biocontrol of anthracnose disease of cowpea. J. Basic Microbiol., 44: 3-9.
CrossRefPubMedDirect Link - Aziz, N.H., M.Z. El-Fouly, A.A. El-Essawy and M.A. Khalaf, 1997. Influence of bean seedling root exudates on the rhizosphere colonization by Trichoderma lignorum for the control of Rhizoctonia solani. Bot. Bull. Acad. Sinica, 38: 33-39.
Direct Link - Chitarra, G.S., T. Abee, F.M. Rombouts, M.A. Posthumus and J. Dijksterhuis, 2004. Germination of Penicillium paneum conidia is regulated by 1-octen-3-ol, a volatile self inhibitor. Applied Environ. Microbiol., 70: 2823-2829.
CrossRefPubMedDirect Link - El-Tarabily, K.A., M.H. Soliman, A.H. Nassar, H.A. Al-Hassani, K. Sivasithamparam, F. McKenna and G.E. Hardy, 2000. Biological control of Sclerotinia minor using a chitinolytic bacterium and actinomycetes. Plant Pathol., 49: 573-583.
Direct Link - Eziashi, E.L., N.U. Uma, A.A. Adekunle and C.E. Airede, 2006. Effect of metabolites produced by Trichoderma species against Ceratocystis paradoxa in culture medium. Afr. J. Biotechnol., 5: 703-706.
Direct Link - Fahima, T. and Y. Henis, 1997. Increasing of Trichoderma hamatum and Talaromyces flavus on the Root of Safe and Unsafe Hostes. In: Biological Control of Soil-Born Plant Pathogens, Hornby, D. (Ed.). (Translated by: Alavi, A. and A. Ahoonmanesh). Research, Education and Extension Organization, Tehran, Iran, pp: 296-322.
- Fridlender, M., J. Inbar and I. Chet, 1993. Biological control of soil-borne plant pathogens by a β-1,3- glucannase-producing Pseudomonas cepacia. Soil Biol. Biochem., 25: 1211-1221.
CrossRef - Guarro, J., J. Gene and A.M. Stchigel, 1999. Developments in fungal taxonomy. Clin. Microbiol. Rev., 12: 454-500.
Direct Link - Hashem, M. and E.H. Ali, 2004. Epicoccum nigrum as biocontrol agent of Pythium damping-off and root-rot of cotton seedlings. Arch. Phytopathol. Plant Prot., 37: 283-297.
CrossRef - Heydari, A., I.J. Misaghi and W.B. McCloskey, 1997. Effects of three soil-applied herbicides on populations of plant disease suppressing bacteria in the cotton rhizosphere. Plant Soil, 195: 75-81.
CrossRefDirect Link - Heydari, A. and I.J. Misaghi, 1998. The impact of herbicides on the incidence and development of Rhizoctonia solani-induced cotton seedling damping-off. Plant Dis., 82: 110-113.
Direct Link - Heydari, A. and I.J. Misaghi, 2003. The role of rhizosphere bacteria in herbicide-mediated increase in Rhizoctonia solani-induced cotton seedling damping-off. Plant Soil, 257: 391-396.
CrossRefDirect Link - Heydari, A., H. Fattahi, H.R. Zamanizadeh, N.H. Zadeh and L. Naraghi, 2004. Investigation on the possibility of using bacterial antagonists for biological control of cotton seedling damping-off in greenhouse. Applied Entomol. Phytopathol., 72: 51-68.
Direct Link - Heydari, A., A. Ahmadi, S. Sarkari, H. Karbalayi Khiavi and M. Delghandi, 2007. Study on the role of common weeds in survival of Verticillium dahliae the causal agent of cotton wilt disease. Pak. J. Biol. Sci., 10: 3910-3914.
CrossRefPubMedDirect Link - Kim, K.K.A. and D.R. Fravel, 1990. Glucose oxidase as the antifungal principle of talaron from Talaromyces flavus. Can. J. Microbiol., 36: 760-764.
PubMedDirect Link - Madi, L., T. Katan, J. Katan and Y. Henis, 1997. Biological control of Sclerotium rolfsii and Verticillium dahliae by Talaromyces flavus is mediated by different mechanisms. Phytopathology, 87: 1054-1060.
PubMedDirect Link - Marois, J.J., D.R. Fravel and G.C. Papavizas, 1984. Ability of Talaromyces flavus to occupy the rhizosphere. Soil Biol. Biochem., 16: 387-390.
Direct Link - Menendez, A.B. and A. Godeas, 1998. Biological control of Sclerotinia sclerotiorum attacking soybean plants: Degradation of the cell wall of this pathogen by Trichoderma harzianum. Mycopathology, 142: 153-160.
CrossRefDirect Link - Mi, S., D.M. Danforth, N.P. Tugwell and M.J. Cohran, 1998. Plant-based injury level for assessing economic thresholds in early season cotton. J. Cot. Sci., 2: 35-52.
Direct Link - Murray, F.R., D.J. Llewellyn, W.J. Peacock and E.S. Dennis, 1997. Isolation of the glucose oxidase gene from Talaromyces flavus and characterization of its role in the biocontrol of Verticillium dahliae. Curr. Genet., 35: 367-375.
Direct Link - Naraghi, L., F. Azad-Disfani and A. Heydari, 2004. The probability of using Talaromyces flavus for biological control of Verticillium wilt of cotton. Proceedings of the 3rd National Conference on the Development in the Application of Biological Products and Optimum Utilization of Chemical Fertilizers and Pesticides in Agriculture, Februrary 21-23, 2004, Seed and Plant Improvement Institute, Karadj, Tehran, Iran, pp: 411.
- Naraghi, L., H. Zareh-Maivan, A. Heydari and H. Afshari-Azad, 2007. Investigation of the effect of heating, vesicular arbuscular mycorrhiza and thermophilic fungus on cotton wilt disease. Pak. J. Biol. Sci., 10: 1596-1603.
CrossRefPubMedDirect Link - Rodriguez, M.A., G. Cabrera and A. Godeas, 2006. Cyclosporine A from a nonpathogenic Fusarium oxysporum suppressing Sclerotinia sclerotiorum. J. Applied Microbiol., 100: 575-586.
CrossRefPubMedDirect Link - Spink, D. and R.C. Rowe, 1989. Evaluation of Talaromyces flavus as a biological control agent against Verticillium dahliae in potato. Plant Dis., 73: 230-236.
CrossRefDirect Link - Tjamos, E.C., 1991. Recovery of olive tree with Verticillium dahliae after individual application of soil solarization in established olive orchards. Plant Dis., 75: 557-562.
CrossRefDirect Link - Tjamos, E.C. and D.R. Fravel, 1995. Detrimental effects of sublethal heating and Talaromyces flavus on microsclerotia of Verticillium dahliae. Phytopathology, 85: 388-392.
Direct Link - Tjamos, E.C. and D.R. Fravel, 1997. Distribution and establishment of the biocontrol fungus Talaromyces flavus in soil and on roots of solanaceous crops. Crop Prot., 16: 135-139.
CrossRef