
Abolfazl Faraji
Gorgan University of Agricultural Sciences and Natural Resources, Gorgan, Iran
Agricultural and Natural Resources Research Center of Golestan, P.O. Box 181,
Asbdavani Street, Gonbad Kavous, Iran
Nasser Latifi
Gorgan University of Agricultural Sciences and Natural Resources, Gorgan, Iran
Afshin Soltani
Gorgan University of Agricultural Sciences and Natural Resources, Gorgan, Iran
Amir Hossain Shirani Rad
Department of Oilseed Crops, Ministry of Jihad-e-Agriculture, Iran
Asian Journal of Plant Sciences
Year: 2008 | Volume: 7 | Issue: 4 | Page No.: 343-351







ABSTRACT
An experiment was conducted in Gonbad, Iran between 2005 and 2007 to study the variation in flower and pod number under high temperature stress and supplemental irrigation. The experiment was a randomized complete block design arranged in split plot. Two cultivars of spring type canola (Hyola401 and RGS003) as subplots and 5 sowing dates as main plots were used. The number of flowers on the main racemes that produced pods was 57 and 49% for Hyola401 and RGS003, respectively. Seed yield was positively related to pods per plant in the two cultivars with 38 and 64% of the variation for Hyola 401 and 71 and 84% of that for RGS003 under irrigation and rainfed conditions, respectively. The relationship between the number of flowers and pods in the main racemes was positively strong with 88 and 72% of the variation for Hyola401 and 88 and 78% of that for RGS003 under irrigated and rainfed conditions, respectively. High temperature; particularly in the late sowing dates accelerated the rate of plant development and reduced the length of the flowering period and also the number of flowers and pods. With a delay in sowing date; flowers and pods per main raceme decreased from 53 to 12 flowers and 34 to 2 pods over two years. Obtained results showed that a great proportion of the variations in flower and pod formation were related to environmental stress during the flowering. Quantitative relationships between flower and pod number with seed yield, crop phenology and temperature pattern during the critical period of flowering could be useful for crop modeling.
PDF Abstract XML References Citation
How to cite this article
Abolfazl Faraji, Nasser Latifi, Afshin Soltani and Amir Hossain Shirani Rad, 2008. Effect of High Temperature Stress and Supplemental Irrigation on Flower and Pod Formation in Two Canola (Brassica napus L.) Cultivars at Mediterranean Climate. Asian Journal of Plant Sciences, 7: 343-351.
DOI: 10.3923/ajps.2008.343.351
URL: https://scialert.net/abstract/?doi=ajps.2008.343.351
DOI: 10.3923/ajps.2008.343.351
URL: https://scialert.net/abstract/?doi=ajps.2008.343.351
INTRODUCTION
The total flower and pod number is a result of the processes that led to their initiation and development. Counts of flowers and pods are useful for breeders and plant physiologists in understanding the reproductive efficiency of the crop (Coffelt et al., 1989). High temperature stress has been shown to cause abortion of flowers in some species, including pea (Pisum sativum L.) (Guilioni et al., 1997), sunflower (Helianthus annuus L.) (Chimenti and Hall, 2001) and canola (Brassica napus L.) (Angadi et al., 2000; Morrison and Stewart, 2002). The adaptation of canola is influenced by tolerance to high temperature, drought stress and by crop management technique which takes the advantage of available optimum environmental conditions during flowering. Canola tolerance to high temperature stress depends on its developmental stage but the reproductive stage is quite susceptible to temperature stress (Angadi et al., 2000). Hall (1992) reported that flowering is the most sensitive stage for temperature stress damage, probably due to vulnerability during pollen development, anthesis and fertilization. In sunflower, Chimenti and Hall (2001) reported that temperatures below 19C or above 30C significantly reduced the number of filled grains. Morrison and Stewart (2002) showed that flowering of canola is inhibited at temperature above 27C while Polowick and Sawhney (1988) reported pod sterility and abortion in canola at 32C. Morrison (1993) reported complete sterile flowers in canola at 27/17C.
Gan et al. (2004) reported that high temperature stress (35/18C) reduced main raceme pods by 75% from the control. There was a significant temperature x cultivar interaction showing effects on pod production under stress treatment. Unlike high temperature stress, water stress had no effect on pod production, either on the main raceme or the branches (Gan et al., 2004). However the direct effects of high temperature stress depend on the crop species and its adaptability (Nuttal et al., 1992; Mahan et al., 1995; Angadi et al., 2000). The contributions of main and branch racemes in pod production depend on environmental conditions. Angadi et al. (2003) reported that under favorable growing conditions, secondary pods contributed between 8-60% of the total pod number. In their study, under the stressful growing conditions, secondary pods contributed 6-38% of the total pod number.
Rao et al. (1992), Guilioni et al. (1997) and Wang et al. (2006) reported that high temperatures accelerate the rate of plant development and causes abortion of flowers with applicable loss in seed yield. Hall (1992) also reported that severe stress reduces the duration of reproductive growth. In canola (Johnston et al., 2002), low water and high temperature stress conditions during flowering and seed filling can result in poor pod formation and seed yield. Heat stress during flowering of canola can prematurely end flowering, resulting in limited seed set. Sowing date can be important to avoid high temperatures at flowering period and at the end of growing season. Therefore there is a need to understand the relationship between high temperature stress and flower production and senescence in canola.
Although the importance of cultivar, sowing date and water stress on canola have been investigated (Brandt and McGregor, 1997; Nielsen, 1997; Chongo and McVetty, 2000; Morrison and Stewart, 2002; Gan et al., 2004; Fan et al., 2005), but there have been no detailed studies on the effects of heat stress and supplemental irrigation on flower, pod production and seed yield. Moreover less is known about the determination of flower and pod number in response to environmental conditions. So, the objective of this study was to investigate flower production and abortion, pod formation and seed yield of canola as affected by supplemental irrigation, sowing date and cultivar in a Mediterranean climate.
MATERIALS AND METHODS
Study was conducted at the Agricultural Research Station of Gonbad (45 m a.s.l., 37 N, 55 E), Golestan province, Iran during 2005-2006 and 2006-2007, under irrigated and rainfed conditions. P and K were applied as basal to supply 50 P2O5 kg ha-1 and 50 K2O kg ha-1 using triple super phosphate and potassium sulfate, respectively. N was applied at 75 kg ha-1 (as urea) with a third applied as basal, a third used to side-dressed at the beginning of stem elongation and the rest applied at the beginning of flowering. The experiment was designed as a randomized complete block design arranged in split plot with 3 replications. Two cultivars of spring canola (namely, Hyola401 and RGS003) were allocated as subplots and five different sowing dates were used as main plots. The sowing dates were 9 Nov., 6 Dec., 5 Jan., 4 Feb. and 6 Mar. in 2005-2006 and 6 Nov., 6 Dec., 5 Jan.,
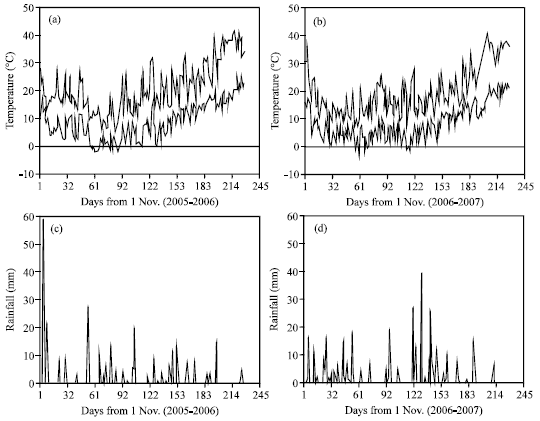 | |
| Fig. 1: | Maximum and minimum daily temperatures (a and b) and daily rainfall (c and d) during the experiment |
4 Feb. and 6 Mar in 2006-2007. The best sowing date for canola in the area is 6-21 Nov., but we planted canola in the late sowing dates to synchronize the reproductive stages of the crop with heat and drought stress periods.
Plots were over planted and after seedling establishment, the plants were thinned to the desired spacing between plants of 5 cm (1000000 plant ha-1). Each subplot consisted of eight rows. Main plots and subplots were 2 and 0.4 m apart, respectively. A 3 m pathway separated replicates. The plots were hand weeded during the season. Two days after beginning of flowering, all of the flowers and buds (each main and branch racemes) were counted and recorded from 10 plants selected at random from within each plot. The plants were tagged and numbered and at the physiological maturity the tagged plants were removed from the plot and the number of pods (complete pods) and incomplete pods (the pods without seed or with abnormal figures) were counted (each main and branch racemes, separately). In each plot, samples during growth season were taken from 10 plants of rows of 2 and 3. At harvested time (2 days after physiological maturity) rows 5, 6 and 7 were cut by hand and air dried. Then the plants were threshed with a stationary combine to determine seed yield. Phonological observations were made on a regular basis with the Harper and Berkenkamp (1975) growth stage key. Maximum and minimum temperatures and rainfall measured at a near-by weather station (Fig. 1). Mean temperatures was calculated for the same growth phases as the sum of the daily temperatures divided by the number of days during the growth phase. Under supplemental irrigation conditions, plots were irrigated at the beginning of stem elongation, flowering and seed filling stages. Two days before irrigation times, soil samples were dried for 24 h at 105C and weighed. Then soil water content was measured and plots were reached to field capacity with irrigation (Zhang et al., 1999). All obtained data were tested by the analysis of variance using SAS (1989) and mean grouping was done according to the LSD (p = 0.05).
RESULTS AND DISCUSSION
Seed yield of canola is determined by the number of flowers per unit land area which produced pods (Morrison and Stewart, 2002). In this study, the combined analysis revealed statistical significant effect of year on flower and pod parameters (data not shown); therefore we analyzed each year data, separately (Table 1). In each year, the number of flowers, pods and pods/flowers ratio were significantly affected by irrigation, sowing dates and cultivars (Table 1). Seed yield was positively related to pods per plant in the two cultivars with 38 and 64% of the variation for Hyola401 and 71 and 84% of that for RGS003 under irrigated and rainfed conditions, respectively (Fig. 2). With increase in number of flowers per plant, the number of pods increased. Pod number produced per plant is mainly set at flowering period, when the number of flowers and pods are determined. The relationship between the number of flowers and pods in the main
| Table 1: | Means of some traits of canola during the experiment |
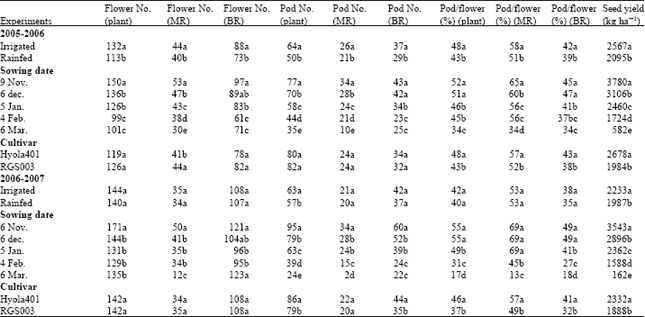 | |
| Means followed by the same letter(s) within each column are not significantly different according to the LSD (p = 0.05). MR: Main Raceme, BR: Branch Racemes | |
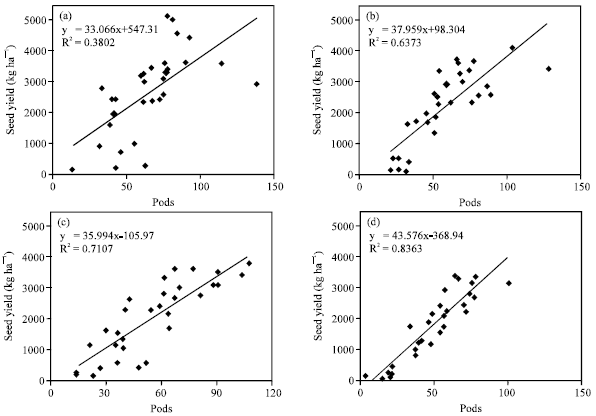 | |
| Fig. 2: | Relationship between seed yield and the number of pods per plant, (a) Hyola401 at irrigated conditions, (b) Hyola401 at rainfed conditions, (c) RGS003 at irrigated conditions and (d) RGS003 at rainfed condition |
sraceme was positively strong with 88 and 72% of the variation for Hyola401 and 88 and 78% of that for RGS003 under irrigated and rainfed conditions, respectively (Fig. 3). This suggests that increase in flower number, particularly in the main raceme under supplemental irrigation, could be an important factor that contributes to increase in pod number in canola.
In the main raceme, the ratio of pods/flowers for Hyola401 and RGS003 was 57 and 52% in 2005-2006 and 57 and 49% in 2006-2007, respectively (Table 1). In branch racemes, the ratio of pods/flowers for Hyola401 and RGS003 was 52 and 39% in 2005-2006 and 41 and 32% in 2006-2007, respectively (Table 1). In each year, the number of flowers that were converted into pods in the main raceme was more than that of branch racemes. McGregor (1981) reported that less than 45% of the flowers formed on B. napus cultivars produced pods. Canola usually produces nearly twice as many flowers as pods. Tayo and Morgan (1975) and McGregor (1981) reported that under typical conditions, canola differentiates more floral primordial than its photosynthetic capacity can support.
The amount of vegetative growth and the weather conditions (temperature, rainfall and radiation) during flowering period determines the formation of buds and flowers and hence the final pod number. In this study, other factors such as days to flowering and temperature during flowering period influenced flower and pod production of canola. There was a positive relationship between the number of flowers per plant with the number of days to flowering (Fig. 4) with 38 and 57% of the variation for Hyola401 and RGS003, respectively. The relationship between the numbers of pods per plant with days to flowering was also very strong with 81 and 76% of the variation for Hyola401 and RGS003, respectively (Fig. 4). This was in agreement with the result of Thurling (1974). There was a strong negative relationship between pod number and the ratio of pods/flowers with mean temperature during the period (Fig. 5) with 84 and 73% of the variation for Hyola401 and 71 and 61% of that for RGS003, respectively. Since optimal temperatures during flowering for B. napus is 20C (Angadi et al., 1999), therefore high temperatures particularly in late sowing dates (Table 2) accelerated the rate of plant development, reduced the length of the flowering period and the number of flowers and pods and thereby seed yield potential as shown by Entz and Flower (1991).
In each year, sowing date significantly affected the number of flowers and pods per plant and the ratio of pods/flowers (Table 1). With delay in sowing, flowers and pods per main raceme decreased, ranging from 53 to 12 flowers and 34 to 2 pods over the two years. In 2006-2007, the number of flowers per branch racemes increased in
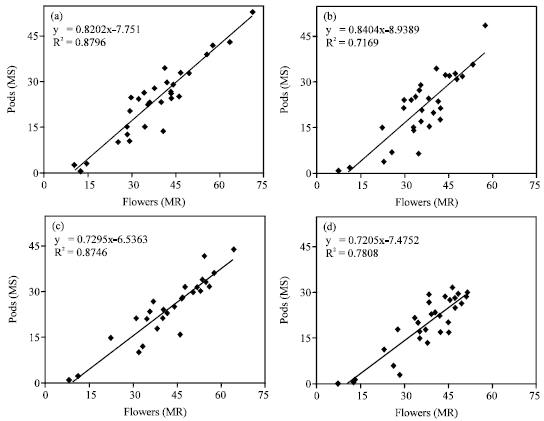 | |
| Fig. 3: | Relationship between the number of flowers and pods in main raceme (MR). (a) Hyola401 at irrigated conditions, (b) Hyola401 at rainfed conditions, (c) RGS003 at irrigated conditions and (d) RGS003 at rainfed conditions |
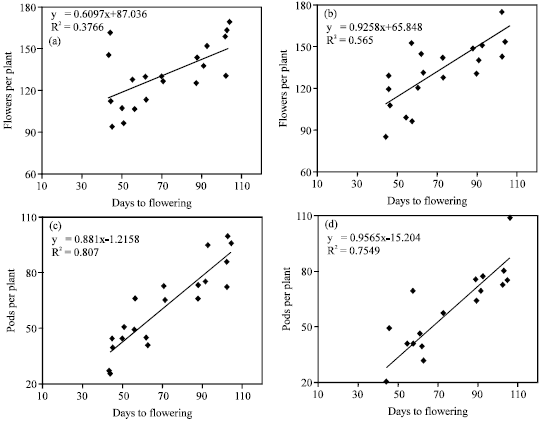 | |
| Fig. 4: | Relationships between days to flowering with the number of flowers per plant (a) Hyola401 and (b) RGS003 and the number of pods per plant (c) Hyola401 and (d) RGS003 for the cultivars |
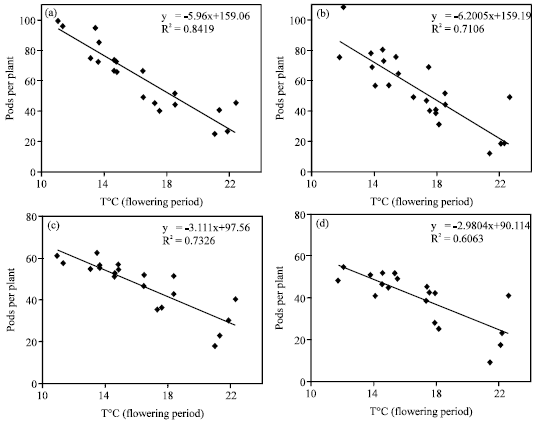 | |
| Fig. 5: | Relationships between pod number per plant (a) Hyola401 and (b) RGS003 and the ratio of pods/flowers per plant (c) Hyola401 and (d) RGS003 with mean temperature during flowering period for the cultivars |
| Table 2: | The mean max. temperature (C) during flowering and seed filling periods |
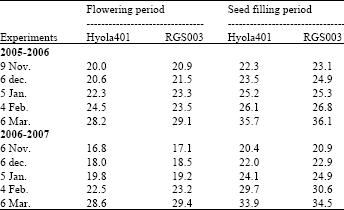 | |
sowing date 5 compared to others, due to good weather conditions and high rainfall in March. (Table 1, Fig. 1). However, the number of flowers in branch racemes that could produce pod decreased significantly due to further severe water and heat stress. Therefore the number of pods per branch racemes in sowing date 5 was the same as sowing date 4 due to severe low ratio of pods/flowers in sowing date 5. Seed yield of canola was found to be closely related to temperature during flowering and early seed development but rainfall from early seed development through seed filling is essential to achieving full yield potential (Johnston et al., 2002). In this experiment, late sowing synchronized critical flowering period with heat stress (Table 2) and thus reduced the duration, flower, pod number and seed yield (Table 1). In North Dakota, the reduction in canola yield with delay in sowing was attributed to fewer pods per plant, leading to a lower harvest index (Johnson et al., 1995). In Gonbad area, with a typical terminal heat and drought stress particularly in late sowing dates (Table 2), heat stress during flowering of canola can prematurely end flowering, resulting in limited flower and pod formation per unit area. However the reproductive period (flowering and seed filling periods) of the early-sown canola is longer than that of the late-sown crop, therefore, contributing to a higher flower and pod number. The chance that rainfed early-sown canola will face heat and drought stress during flowering and seed filling periods was not as high as they would be if the crop was sown late.
In each year, with delay in sowing, the ratio of pods/flowers decreased (Table 1) and that was due to increase in terminal heat stress, though the rate of decrease was not the same. This was confirmed by the strong positive relationship between incomplete pods and flowers in the main raceme in sowing date 5, which accounted for 84 and 89% of the variation for Hyola401 and RGS003, respectively (Fig. 6). There was not any significant relationship between incomplete pods and
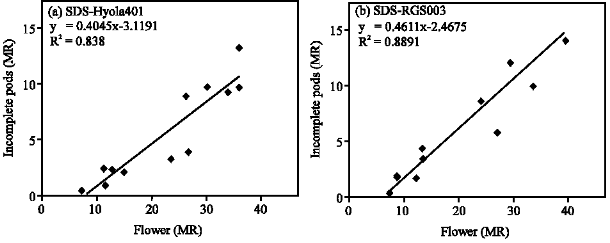 | |
| Fig. 6: | Relationship between flower number per plant and incomplete pods in Main Raceme (MR) at sowing date 5 (SD5) for the cultivars |
flowers in the main raceme in early sowing dates. The rate of decrease in the ratio of pods/flowers in 2006-2007 was more than that of 2005-2006 ranging from 65-34% in main raceme, 47-34% in branch racemes and 52-34% in total plant in 2005-2006 and 69-13% in main raceme, 49-18% in branch racemes and 55-17% in total plant in 2006-2007 (Table 1). With delay in sowing, the rate of decrease in the ratio of pods/flowers in the main raceme was more than that of branch racemes due to decrease or/even increase in flowers per plant in branch racemes compared to the main raceme.
In 2005-2006, supplemental irrigation was significantly increased the number of flowers and the ratio of pods/flowers (both main and branch racemes), compared to rainfed conditions, but there was no significant difference between flower number and pods/flowers ratio in 2006-2007 (Table 1). Under supplemental irrigation conditions, the ratio of pods/flowers in the main raceme, branch racemes and total plant was 58, 42 and 48% in 2005-2006 and 53, 32 and 37% in 2006-2007, respectively (Table 1). Under rainfed conditions, the ratio of pods/flowers in the main raceme, branch racemes and total plant was 51, 39 and 43% in 2005-2006 and 49, 32 and 37% in 2006-2007 (Table 1). It appeared that the benefit of supplemental irrigation in 2005-2006 was more than that of 2006-2007. The cumulative rainfall from October to June in 2005-2006 and 2006-2007 was 393 and 448 mm, respectively (Fig. 1) and as such, supplemental irrigation amount was higher in the first year of the experiment (data not shown). When the amount of rainfall from March to May between the two years were compared and, flowering and seed filling periods of canola in the area with normal sowing dates, the difference was higher. The cumulative rainfall from March to May in 2005-2006 and 2006-2007 was 98 and 229 mm, respectively (Fig. 1). Therefore water stress was more at the time of flowering and seed filling periods in 2005-2006 than 2006-2007, providing a better opportunity for supplemental irrigation to influence canola flower number and pods/flower ratio in 2005-2006 (Table 1). This was confirmed with the result of seed yield in the experiment. However seed yield of canola under irrigation conditions, sowing dates and cultivars in 2005-2006 (2331 kg ha-1) was more than that of in 2006-2007 (2110 kg ha-1), due to better weather conditions and distribution of rainfall during crop growth season in 2005-2006 (Table 1, Fig. 1).
The mean annual rainfall of Gonbad area is about 450 mm. Because approximately there is no enough rainfall in April till May, coinciding with the seed filling period of canola in the area, occurrence of drought and crop water stress is common. The heat (Table 2) and drought stress that normally occurs in the area significantly lowers the seed yield of canola. Increased production, as a result, depends to a greater extent on further improvement of soil water availability. Therefore drought effects can be minimized by management practices such as supplemental irrigation (Oweis et al., 2004; Fan et al., 2005). Brandt and McGregor (1997) reported that for each millimeter increase in rainfall, there was a corresponding seed yield increase. The number of flowers (each main and branch racemes) in the two cultivars over years, sowing dates and irrigation conditions were approximately the same, but in both years, the ratio of pods/flowers of Hyola401 was more than that of RGS003 (Table 1). The mean 2 year of the ratio of pods/flowers for Hyola401 and RGS003 was 57 and 50% in main raceme, 42 and 35% in branch racemes and 47 and 40% in total plant, respectively (Table 1).
CONCLUSION
Pod formation and seed yield of canola in semiarid environments can be increased by minimizing the crop exposure to high temperatures and water stress that often occurs during the growing season. In this study, earlier sowing more closely matched incoming rainfall and reduced terminal temperature stress that increased the number of flowers and pods. Delay in sowing led to more rapid development of the crop, decreased days to flowering, flowering period and flower and pod number per plant. Sowing date and supplemental irrigation were very important management tools in minimizing the negative impact of high temperature and water stress during the reproductive stage in canola. In this study, a great proportion of the variation in flower, pod formation and seed yield in canola were related to environmental conditions during the critical period of flowering. Supplemental irrigation was a viable option that could be used to increase and stabilize canola pod number and seed yield.
REFERENCES
- Angadi, S.V., H.W. Cutforth, P.R. Miller, B.G. McConkey, M.H. Entz, A. Brandt and K.M. Olkmar, 2000. Response of three Brassica species to high temperature stress during reproductive growth. Can. J. Plant Sci., 80: 693-701.
Direct Link - Angadi, S.V., H.W. Cutforth, B.G. McConkey and Y. Gan, 2003. Yield adjustment by canola grown at different plant populations under semiarid conditions. Crop Sci., 43: 1358-1366.
Direct Link - Chimenti, C.A. and A.J. Hall, 2001. Grain number responses to temperature during floret differentiation in sunflower. Field Crops Res., 72: 177-184.
CrossRef - Chongo, G. and P.B.E. McVetty, 2001. Relationship of physiological characters to yield parameters in oilseed rape (B. napus). Can. J. Plant Sci., 81: 1-6.
Direct Link - Coffelt, T.A., M.L. Seaton and S.W. VanScoyoc, 1989. Reproductive efficiency of 14 Virginia type peanut cultivars. Crop Sci., 29: 1217-1220.
Direct Link - Entz, M.H. and D.B. Fowler, 1991. Agronomic performance of winter versus spring wheat. Agron. J., 83: 527-532.
Direct Link - Fan, T., B.A. Stewart, W.A. Payne, Y. Wang, S. Song, J. Luo and C.A. Robinson, 2005. Supplemental irrigation and water-yield relationships for plasticulture crops in the Loess Plateau of China. Agron. J., 97: 177-188.
Direct Link - Gan, Y., S.V. Angadi, H. Cutforth, D. Potts, V.V. Angadi and C.L. McDonald, 2004. Canola and mustard response to short periods of temperature and water stress at different developmental stages. Can. J. Plant Sci., 84: 697-704.
Direct Link - Guilioni, L., J. Wery and F. Tardieu, 1997. Heat stress-induced abortion of buds and flowers in Pea: Is sensitivity linked to organ age or to relations between reproductive organs?. Ann. Bot. London, 80: 159-168.
Direct Link - Harper, F.R. and B. Berkenkamp, 1975. Revised growth-stage key for Brassica campestris and B. napus. Can. J. Plant Sci., 55: 657-658.
Direct Link - Johnson, B.L., K.R. McKay, A.A. Schneiter, B.K. Hanson and B.G. Schatz, 1995. Influence of planting date on canola and crambe production. J. Prod. Agric., 8: 594-599.
Direct Link - Johnston, A.M., D.L. Tanaka, P.R. Miller, S.A. Brandt, D.C. Nielsen, G.P. Lafond and N.R. Riveland, 2002. Oilseed crops for semiarid cropping systems in the Northern great plains. Agron. J., 94: 231-240.
Direct Link - Mahan, J.R., B.L. McMicheal and D.F. Wanjura, 1995. Methods for reducing the adverse effects of temperature stress on plants: A review. Environ. Exp. Bot., 35: 251-258.
CrossRef - McGregor, D.I., 1981. Pattern of flower and pod development in rapeseed. Can. J. Plant Sci., 61: 275-282.
Direct Link - Morrison, M.J., 1993. Heat stress during reproduction in summer rape. Can. J. Bot., 71: 303-308.
CrossRef - Morrison, M.J. and D.W. Stewart, 2002. Heat stress during flowering in summer Brassica. Crop Sci., 42: 797-803.
Direct Link - Nielsen, D.C., 1997. Water use and yield of canola under dry land conditions in the Central Great Plains. J. Prod. Agric., 10: 307-313.
Direct Link - Nuttal, W.F., A.P. Moulin and L.J. Townley-Smith, 1992. Yield response of canola to nitrogen, phosphorus, precipitation and temperature. Agron. J., 84: 765-768.
CrossRef - Oweis, T., A. Hachum and M. Pala, 2004. Lentil production under supplemental irrigation in a Mediterranean environment. Agric. Water Manage., 68: 251-265.
CrossRef - Polowick, P.L. and V.K. Sawhney, 1988. High temperature induced male and female sterility in canola (B. napus L.). Ann. Bot. (London), 62: 83-86.
Direct Link - Rao, G.U., A. Jain and K.T. Shivanna, 1997. Effects of high temperature stress on Brassica pollen: Viability, germination and ability to set fruits and seeds. Ann. Bot. (London), 69: 193-198.
Direct Link - Thurling, N., 1974. Morphophysiological determinants of yield in rapeseed (B. campestris and B. napus). II. yield components. Aust. J. Agric. Res., 25: 711-721.
CrossRef - Wang, J., Y.T. Gan, F. Clarke and C.L. McDonald, 2006. Response of chickpea yield to high temperature stress during reproductive development. Crop Sci., 46: 2171-2178.
Direct Link - Zhang, H.P., X.Y. Wang, M.Z. You and C.M. Liu, 1999. Water-yield relations and water use efficiency of winter wheat in the North China plain. Irrig. Sci., 19: 37-45.
CrossRef