
Juthaporn Khampila
Not Available
Piyada Theerakulpisut
Not Available
Kamol Lertrat
Not Available
Weerasak Saksirirat
Not Available
Jirawat Sanitchon
Not Available
Nooduan Muangsan
Not Available
Asian Journal of Plant Sciences
Year: 2008 | Volume: 7 | Issue: 1 | Page No.: 18-21







ABSTRACT
A F2 population from a cross of the waxy corn inbreds 209W and 241 W was used to determine random amplified polymorphic DNA (RAPD) markers linked to Northern Corn Leaf Blight (NCLB) resistance via Bulked Segregant Analysis (BSA). Resistant and susceptible DNA bulks were constructed using the segregating F2 plants based on phenotypic reaction against NCLB infection. In total, two hundred and twenty two decamer primers were used to identify three polymorphic bands observed between the bulks. The primers OPE02, OPJ18 and OPX04, generated the polymorphic DNA fragments of approximately 1200, 650 and 500 bp, respectively. These RAPD bands were present in only in NCLB-resistant bulk and 241W resistant parent. The study revealed that RAPD markers associated to NCLB resistance is potentially useful for the identification of genotyped individuals carrying NCLB resistant traits in breeding programs.
PDF Abstract XML References Citation
How to cite this article
Juthaporn Khampila, Piyada Theerakulpisut, Kamol Lertrat, Weerasak Saksirirat, Jirawat Sanitchon and Nooduan Muangsan, 2008. Identification of RAPD Markers for Northern Corn Leaf Blight Resistance in Waxy Corn (Zea mays var. ceratina). Asian Journal of Plant Sciences, 7: 18-21.
DOI: 10.3923/ajps.2008.18.21
URL: https://scialert.net/abstract/?doi=ajps.2008.18.21
DOI: 10.3923/ajps.2008.18.21
URL: https://scialert.net/abstract/?doi=ajps.2008.18.21
INTRODUCTION
Exserohilum turcicum (Pass.) K.J. Leonard and E.G. Suggs causes Northern Corn Leaf Blight (NCLB), a severe and widespread foliar wilt disease of maize (Zea mays L.) (Leonard et al., 1989). NCLB causes substantial crop losses in most of the major corn producing areas throughout the world, particularly in mid-altitude and highland regions of the tropics and subtropics (Smith and Kinsey, 1993). Symptoms can range from small cigar-shaped lesions to complete destruction of the foliage. Grain yield losses can exceed 50% in susceptible maize cultivars if infection occurs before flowering (Tefferi et al., 1996). To prevent severe yield losses, the use of resistant cultivars is currently the most effective and common means of disease control and also very important due to concern about pesticides and the environment. Breeding for NCLB resistant maize cultivars by conventional means is considered the most effective and feasible method to overcome yield losses due to NCLB. However, the conventional breeding is laborious, time-consuming and dependent on environmental conditions. The use of molecular markers is an efficient alternative to the tedious work of phenotype evaluation for NCLB resistance and allows for an efficient selection of NCLB resistance gene (s). Molecular markers can also accelerate selection and eliminate the effects of environmental variation during selection (Malyshev and Kartel, 1997).
Random Amplified Polymorphic DNA (RAPD) analysis has been successfully used to identify DNA polymorphism linked to many important traits such as disease resistant genes in sugar pine (Devey et al., 1995), tobacco mosaic virus resistant gene in tomato (Young et al., 1998), downy mildew resistant gene in sunflower (Brahm et al., 2000), angular leaf spot disease resistant gene in common bean (Ferreira et al., 2000), soybean mosaic virus resistant gene in soybean (Zheng et al., 2001), powdery mildew resistant gene in grapes (Dalbo et al., 2001). RAPD is a simple and inexpensive technique compared to Restriction Fragment Length Polymorphism (RFLP). RAPD markers are also more-rapidly and more-easily detected than RFLP markers (Welsh and McClelland, 1990; Williams et al., 1990). Michelmore et al. (1991) suggested an alternative method called Bulked Segregant Analysis (BSA) to find RAPD markers linked to the trait of interest, such as resistant to NCLB disease. In the present studies RAPD analysis was used to identify DNA marker (s) linked to NCLB resistance in waxy corn using bulked segregant analysis in F2 population.
MATERIALS AND METHODS
A 217 F2 population from a cross between a highly susceptible inbred 209W line with a resistant inbred 241W line was grown along with the parents in the field area of the Department of Plant Science and Agricultural Resources, Faculty of Agriculture, Khon Kaen University, Thailand during the normal growing season of maize of the year 2005. The resistant inbred 241W was originally derived from derived from the open-pollinated variety Sumlee Esan, while the susceptible inbred 209W was originally derived from a cross between the CK8-F2 and KSC-F2 (Thoungnarin et al., 2005). The F2 population was screened for NCLB resistance trait. Severity of NCLB was assessed, based on the percentage of the total leaf area affected using a slightly modified version of the standard protocol of Elliot and Jenkins (1946). Parent genotypes were screened for polymorphism using 222 RAPD primers including 11 primer sets from Operon technologies, Alameda, CA (Sets A, B, C, E, F, G, H, J, W, X and Y), UBC244 and UBC9. PCR amplifications were carried out according to the method described in Williams et al. (1990). For the bulks, resistant and susceptible bulks were prepared from F2 individuals by pooling aliquots containing equivalent amounts of total DNA from each of the fourteen extremely resistant and fourteen extremely susceptible F2 progenies. DNA was extracted separately from each individual of the progeny. Genomic DNA of young leaves before inoculation was extracted by the method of Doyle and Doyle (1987) with minor modifications and adapted to small tissue quantities (Hormaza, 1999). The PCR reaction volume was 25 μL and contained: 1X PCR buffer, 2.0 mM MgCl2, 300 μM of dNTPs, 0.4 μM of primer, 0.7 U of Tag DNA polymerase (Promega®, Madison, Wisconsin) and 10 ng of genomic DNA. DNA amplification was conducted on a programmable thermalcycler (Hybaid®, USA). Template DNA was initially denatured at 94C for 2 min, followed by 47 cycles of PCR amplification using the following parameters: 1 min denaturation at 94C, 1.45 min annealing at 38C and 2 min primer extension at 72C. A final 7 min incubation at 72C was allowed for completion of primer extension. PCR products were electrophoretically resolved on 1.5% agarose gels with 1X TBE (40 mM Tris-borate, 1 mM EDTA) containing 0.5 g mL-1 ethidium bromide and detected on a UV transilluminator.
Bulked segregant analysis: The method involves comparing two pooled DNA samples from individuals of two extreme phenotypes from a segregating population. Within each pool or bulk, the individuals are identical for the trait of interest but are arbitrary for all other traits.
Therefore, the two resultant bulked DNA samples differ genetically only in the selected region and are seemingly heterozygous and monomorphic for all other regions. Two DNA pools contrasting for the trait of interest are analyzed to identify markers that distinguish them. Markers that are polymorphic between the pools will be genetically linked to the loci determining the trait used to construct the pools (Michelmore et al., 1991). Two bulked DNA samples were generated from the F2 segregating population. One bulk consisted of equal amounts of DNA of fourteen NCLB resistant F2 plants and the other was similarly formed from DNA of fourteen F2 plants with susceptible to NCLB following the procedure giving by Michelmore et al. (1991). PCR reactions were carried out on the bulks and parental DNA samples using RAPD primers that were polymorphic between the parents.
RESULTS AND DISCUSSION
Out of 222 arbitrary decamer primers screened for polymorphisms between 209W susceptible and 241W resistant parents, 63 RAPD primers (28.38%) that gave polymorphic bands between the parent genotypes were identified. Moreover, 51 oligonucleotide 10-mer primers were not obtained any DNA bands and 14 RAPD primers gave very weak DNA profiles, while 94 RAPD primers gave the monomorphic bands when the 222 RAPD primers were used to study the polymorphism between the parents. The total 1,069 bands were amplified using the 222 RAPD primers. The number of RAPD fragments that were amplified ranged from 2 to 11 and the sizes ranged from about 200 to 1,300 bp. Of these 222 RAPD primers, sixteen primers which produced single, strong polymorphic bands that were present in only 241W resistant parent, but absent in 209W susceptible parent, were selected for screening DNA bulks and their parental DNA.
In RAPD, an often occurring artifact on agarose gels is a primer and the target sequence not matching 100% would be less so amplification would be quantitatively less from these loci resulting in faint bands. Extremely bright RAPD bands observed in the present studies may be the result of amplification from sequences of high copy number in the genome. The amount of amplification product of a sequence of high copy number is expected to be greater compared to that of low copy number which would result in a very bright band (Malik, 1995). However, a few bright bands split into sub-bands when ran on relatively high concentration agarose gel (2.5%). This suggested that they were a mixture of fragments in a small range of sizes (Amir et al., 2002).
A total of sixteen RAPD primers were used for screening the bulks containing bulked DNA from fourteen plants each from the resistant and susceptible F2 plants as described. The sixteen RAPD primers that generated polymorphic DNA fragments between parents were analyzed and the average number of bands per primer was again between 4 and 5. Screening of bulks and parental lines with the primers showed amplification products ranging in size between 300 to 1,300 bp. The primers OPE02 (5´-GGTGCGGGAA-3´), OPJ18 (5´-TGGTCGCAGA-3´) and OPX04 (5´-CCGCTACCGA-3´), generated the DNA fragments of approximately 1200, 650 and 500 bp, respectively (Fig. 1). These RAPD markers were present only in NCLB-resistant bulk and 241W parent, but were missing in NCLB-susceptible bulk and 209W parent and the markers were reproducible. These three RAPD primers generated polymorphic fragments that are associated to the NCLB resistance phenotypes.
Random primers used in RAPD analysis usually anneal with multiple sites in different regions of the genome so that several genetic loci are amplified and also the markers are inherited as dominant genetic markers. This limits the application of this marker type, particularly in cases where one would like to distinguish homozygous from heterozygous genotypes. The PCR amplification that generates RAPD fragments of interest is sensitive to specific reaction conditions. Moreover, poor reproducibility can occur in RAPD analysis. A different population may produce different amplification profiles using the same primers. Nevertheless, the enormous attraction of RAPD marker is that the technique is quick, simple, uses small amounts of DNA, sample throughput can be high and the procedure is automatable (Welsh and McClelland, 1990). There is also no requirement for DNA probes or sequence information for primer design when one uses RAPDs.
These studies suggested that RAPD technology has great potential in finding DNA marker (s) for practical breeding programs. Some earlier workers have demonstrated the use of RAPD in practical plant breeding and DNA marker assisted back crossing (Stuber, 1995). RAPD markers linked to rust resistance in barley have been used to carry out successful marker assisted selection in an F2 population (Poulsen et al., 1995). Speed and the efficiency of the crop improvement programs can be enhanced significantly by using marker assisted selection and it also allows consistent progress in the advancement of selected materials. It is important especially for those characters which are highly dependent upon the environment for expression such as drought resistance (Malik, 1995).
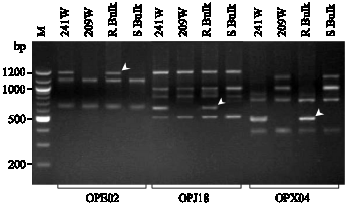 | |
| Fig. 1: | PCR banding patterns generated by primers OPE02, OPJ18 and OPX04 with NCLB resistant parent 241W NCLB susceptible parent 209W, R-bulk the NCLB resistant bulk, S-bulk the NCLB susceptible bulk and M is 100 bp DNA ladder. Arrows indicate the polymorphic DNA band corresponding to NCLB resistance |
Using a method inspired by BSA, we were able to identify three RAPD markers associated to NCLB resistance phenotype in waxy corn, Z. mays var. ceratina. From a total of 1,069 fragments, only three (0.28%) were linked to the NCLB resistance. The RAPD markers should be useful for marker-assisted selection. Present results support the idea that BSA can provide fast detection of molecular markers linked to genes of interest. Traditional methods of handling breeding populations take very long time for advancement to a desired stage. Expenditure using conventional means of breeding such as management and the labor costs of experiments may be higher compared to using marker assisted selection. It is concluded that RAPD technique has great potential in plant breeding.
Further work will involve the conversion of the three associated markers, described in this study to a Sequence Characterized Amplified Region (SCARs) in order to simplify their use in maize breeding programs. And also the construction of the RAPD markers combined with simple sequence repeat (SSR) markers on the genetic map of maize for detecting the Quantitative Trait Loci (QTLs) linked to NCLB resistance genes.
ACKNOWLEDGMENTS
This research was funded by the University Staff Development Program, Mahasarakham University, Thailand and the Agricultural Biotechnology Project, Faculty of Agriculture, Khon Kaen University under the sponsorship of the Asian Development Bank (ADB) Loan and the Thai Government.
REFERENCES
- Amir, A.H., T.A. Malik, H. Raza and M. Rahman, 2002. DNA marker studies for leaf nectaries in upland cotton. Asian J. Plant Sci., 1: 395-396.
CrossRefDirect Link - Brahm, L., R. Thomas and F. Wolfgang, 2000. PCR-based markers facilitating marker assisted selection in sunflower for resistance to downy mildew. Crop Sci., 40: 676-682.
Direct Link - Dalbo, M.A., G.N. Ye, N.F. Weeden, W.F. Wilcox and B.I. Reisch, 2001. Marker assisted selection for powdery mildew resistance in grapes. J. Am. Soc. Hortic. Sci., 126: 83-89.
Direct Link - Devey, E.M., A. Delfino-Mix, B.B. Kinloch and B.D. Neale, 1995. Random amplified polymorphic DNA markers tightly linked to a gene for resistance to white pine blister rust in sugar pine. Proc. Natl. Acad. Sci. USA., 92: 2066-2070.
PubMedDirect Link - Doyle, J.J. and J.L. Doyle, 1987. A rapid DNA isolation procedure for small quantities of fresh leaf tissue. Phytochem. Bull., 19: 11-15.
Direct Link - Ferreira, C.F., A. Borem, G.A. Carvalho, S. Nietsche, T.J. Paula, E.G. Barros and M.A. Moreira, 2000. Inheritance of angular leaf spot resistance in common bean and identification of a RAPD marker linked to a resistance gene. Crop Sci., 40: 1130-1133.
Direct Link - Hormaza, J.I., 1999. Early selection in cherry combining RAPDs with embryo culture. Sci. Hortic., 79: 121-126.
CrossRefDirect Link - Leonard, K.J., Y. Levy and D.R. Smith, 1989. Proposed nomenclature for pathogenic races of Exserohilum turcicum on corn. Plant Dis., 73: 776-777.
Direct Link - Malyshev, S.V. and N.A. Kartel, 1997. Molecular markers in mapping of plant genomes. J. Mol. Biol., 31: 163-171.
Direct Link - Michelmore, R.W., I. Paran and R.V. Kesseli, 1991. Identification of markers linked to disease-resistance genes by bulked segregant analysis: A rapid method to detect markers in specific genomic regions by using segregating populations. Proc. Nat. Acad. Sci. USA., 88: 9828-9832.
PubMedDirect Link - Poulsen, D.M.E., R.J. Henry, R.P. Johnston, J.A.G. Irwin and R.G. Rees, 1995. The use of bulk segregant analysis to identify a RAPD marker linked to leaf rust resistance in barley. Theor. Applied Genet., 91: 270-273.
CrossRefDirect Link - Stuber, C.W., 1995. Mapping and manipulating quantitative traits in maize. Trends-Genet. Camb., 11: 477-481.
CrossRefDirect Link - Assefa, T., H. Mengistu and H.G. Welz, 1996. Evaluation of maize germplasm for resistance to turcicum leaf blight in Bako area, western Ethiopia. Western Ethiopia Afr. J. Plant Prot., 6: 75-82.
Direct Link - Welsh, J. and M. McClelland, 1990. Fingerprinting genomes using PCR with arbitrary primers. Nucleic Acids Res., 18: 7213-7218.
CrossRefPubMedDirect Link - Williams, J.G.K., A.R. Kubelik, K.J. Livak, J.A. Rafalski and S.V. Tingey, 1990. DNA polymorphisms amplified by arbitrary primers are useful as genetic markers. Nucleic Acids Res., 18: 6531-6535.
CrossRefPubMedDirect Link - Young, N., D. Zamir, M.W. Ganal and S.D. Tanksley, 1988. Use of isogenic lines and simultaneous probing to identify DNA markers tightly linked to the Tm-2a gene in tomato. Genetics, 120: 579-585.
PubMedDirect Link - Zheng, C., R. Chang, L. Qiu, M. Zheng, Z. Chang and J. Qiu, 2001. Inheritance of resistance to SMV3 and identification of RAPD marker linked to the resistance gene in soybean. Sci. Agric. Sinica, 34: 14-18.
Direct Link