
Preecha Prathepha
Walai Rukhavej Botanical Research Institute (WRBRI), Mahasarakham University,Khamriang District, Maha Sarakham Province, 44150, Thailand
Asian Journal of Plant Sciences
Year: 2008 | Volume: 7 | Issue: 2 | Page No.: 156-162







ABSTRACT
Amylose content, an important determinant of rice starch quality, is primarily controlled by the Waxy (wx) gene encoding Granule-Bound Starch Synthase (GBSS). Oryza rufipogon, known as the ancestor of Asian cultivated rice (O. sativa L.), is the most important wild relative for rice improvement. To obtain basic information on the diversity of wx microsatellite alleles and on the relationship between wx microsatellite alleles and amylose classes, 212 strains of wild rice were collected and analyzed, using the PCR-RFLP method and DNA sequencing. Seven wx microsatellite alleles, (CT)n repeats, (n = 9, 10, 11, 12, 16, 17 and 18) were found in 16 natural populations. Among them, the (CT)n repeats n = 9, 10, 11, 12 and 16 had the sequence G at the donor splice site and were associated with intermediate to high amylose wild rice strains. The (CT)18 allele, carrying the sequence T at this position, [(CT)18/T], was found only in wild rice strains with low amylose content. The (CT)17 allele was polymorphic for the G-T mutation at the donor splice site of intron 1 of the wx locus. Rice strains containing the (CT)17 allele with G at the donor splice site, [(CT)17/G], showed intermediate to high amylose content, while the allele with the sequence T at this position, [(CT)17/T], was found in rice strains with low amylose content. Variation of DNA-based genetic marker known as the waxy microsatellite corresponds to the amylose class of wild rice is important, as it should be useful in breeding programs designed to produce rice of desired grain quality and used as DNA marker in gene flow studies between cultivated and wild rice.
PDF Abstract XML References Citation
How to cite this article
Preecha Prathepha, 2008. Variation of the Waxy Microsatellite Allele and its Relation to Amylose Content in Wild Rice (Oryza rufipogon Griff.). Asian Journal of Plant Sciences, 7: 156-162.
DOI: 10.3923/ajps.2008.156.162
URL: https://scialert.net/abstract/?doi=ajps.2008.156.162
DOI: 10.3923/ajps.2008.156.162
URL: https://scialert.net/abstract/?doi=ajps.2008.156.162
INTRODUCTION
O. rufipogon is a common perennial wild rice widely distributed in the tropics and subtropics of Asia (Vaughan, 1994). This species is native to all regions of Thailand and grows predominantly in ponds and roadside ditches (Chitrakon, 1995). It is recognized as the progenitor of the Asian cultivated rice (O. sativa) and plays an important role for continued availability and sustainable use of the valuable genetic resources. Therefore, investigation for useful genes in its germplasm has been carried out and some of these genes have successfully been transferred to varieties of cultivated rice reviewed by Song et al. (2005). From conservation point of view, genetic variation and genetic structure of O. rufipogon populations in China were studied by several authors, such as Cai et al.(1998), Gao et al. (2000, 2002), Gao (2004), Song et al. (2003a) and Zhou et al. (2003). In Thailand, the results of Akimoto et al. (1999) suggested thatO. rufipogon showed rapid genetic erosion and action for its conservation in natural populations was urgently needed.
Starch is mainly composed of two types of glucan polymer, amylose and amylopectin (Smith et al., 1997). The gene, which controls amylose synthesis in endosperm and pollen, is called the Waxy (wx) gene. In rice, the amylose content is the most important factor determining rice grain quality for eating and processing (Webb, 1991). Amylose content in rice has been reported to vary from 0 to 33%. Amylose contents of 0-2, 5-12, 12-20, 20-25 and 25-33% have been specified as waxy, very low, low, intermediate and high amylose, respectively (Juliano, 1992). Rice grains with low amylose levels are associated with tender, cohesive, glossy grains when cooked, while grains with higher amylose levels appear dry, fluffy and separated as cooked rice.
Bligh et al. (1995) reported first a polymorphic microsatellite (CT repeats) in the Wx gene. Later, Ayres et al. (1997) reported seven wx microsatellite alleles (n = 8, 11, 14, 17, 18, 19 and 20) and these were found to be correlated with variation in amylose content in 89 U.S. non-glutinous rice cultivars. Six wx microsatellite alleles (n = 10, 11, 14, 17, 18 and 20) were found in the non-glutinous rice samples showing correlation with amylose content (Bergman et al., 2001). Bao et al. (2002) described four (n = 16, 17, 18 and 19) wx microsatellite alleles in Chinese glutinous rice cultivars. Recently, Prathepha and Baimai (2004) reported that four wx microsatellite alleles (n = 16, 17, 18 and 19) were found in glutinous Thai rice, whereas non-glutinous Thai rice has five wx microsatellite alleles (n = 11, 16, 17, 18 and 19). The (CT)11 allele was found only in intermediate and high amylose rice cultivars.
There are several reports in which this gene has been well studied in cultivated rice with respect to both, allele variation and expression (Bligh et al., 1998; Cai et al., 1998; Hirano et al., 1998; Isshiki et al., 1998, 2001). Obviously, the studies of the wx gene were mostly limited to Asian cultivated rice, so far none were carried out on its ancestor, O. rufipogon. The objectives of this research were to explore the distribution of the wx microsatellite alleles. In populations of O. rufipogon in Thailand and Lao PDR and to determine the relation between the wx microsatellite allele and amylose class in wild rice species.
MATERIALS AND METHODS
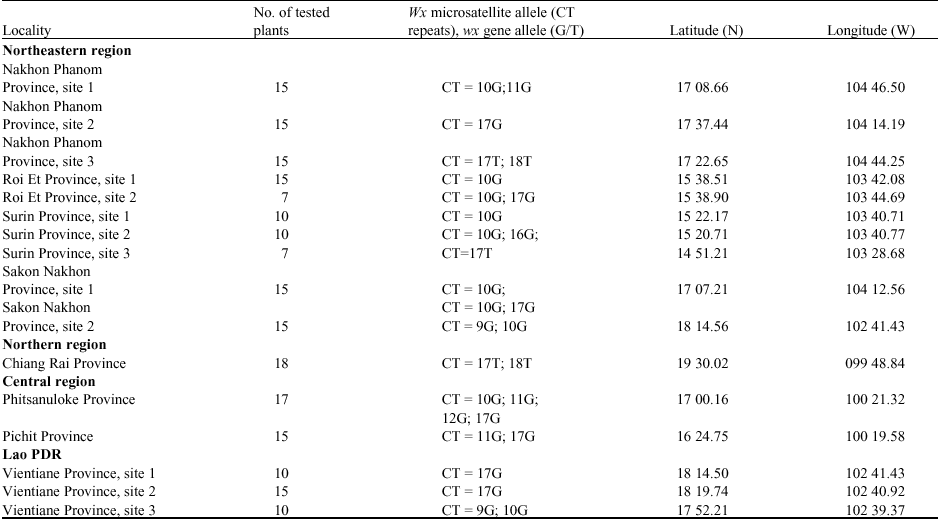
Plant materials: A total of 212 strains of O. rufipogon, including 35 strains collected from Lao PDR and 177 strains from all regions of Thailand were used in this study. The collection had made between October to December 2005-2006. Their collection sites were shown in Table 1. The precise collection sites were documented with Global Positioning System (GPS) from GARMIN (iQue 3600), Garmin Co. Ltd.). Young leaves and seeds were collected individually from at least 10 individuals per population. Because O. rufipogon is a spreading perennial plant with clonal growth. Samples were randomly collected at an interval of at least 5 m to prevent collection of multiple samples from a single genet, as suggested by Gao (2004).
DNA extraction: Total genomic DNA was extracted from fresh leaves following the procedure of DNA isolation kit (DNA Trap, DNA Technology Laboratory, National Center for Genetic Engineering and Biotechnology at Kasetsart University, Kamphangsaen, Nakorn Pathom, Thailand). Fresh young leaves from rice plants were ground in a mortar containing 1 mL of extraction buffer by using a pestle and then the extraction solution was transferred to a microtube. The solutions were mixed and incubated at 65°C for 10 min. The samples were put on ice for 5 min and Neutralizer solution was added. Tubes were then centrifuged at 14,000 x g for 5 min. The supernatant was transferred to a new microtube by using micropipette and 500 μL of the Trapping buffer was added to the samples. Tubes were vortexed briefly and then centrifuged at 14,000 x g for 3-5 sec. The supernatant was discarded. Washing buffer II was added to each tube and the same procedure was performed as with Washing buffer I. The pellets were allowed to air-dry at room temperature, re-suspended in 100 μL of the Elution buffer and incubated at 65°C for 5 min. and centrifuged at 14,000 x g for 3-5 sec. The supernatant was transferred to new microtubes and stored at -20°C until use.
| Table 1: | Location of the populations of O. rufipogon in the present work and wx microsatellite allele at the Waxy gene |
 | |
The wx microsatellite allele and Wx gene allele analysis using PCR assay and DNA sequencing: PCR amplification of the wx microsatellite was performed using the primer pairs : Waxy-F (5´-ACCATTCCTTCAGTTT-3´); Waxy-R (5´-TAGCATGTATGAGACTACTTGTAA-3´). These primer pairs have been reported previously (Prathepha and Baimai, 2004). The PCR amplified region flanks the beginning of exon 1 and the beginning of the intron 1. The PCR reaction was performed in a 20 μL reaction mixture, containing 2 μL of DNA solution, 10 pmol of each of the primer pairs, 1.5 mM MgCl2, 2 units Taq polymerase (Promega), 0.1 mM dNTPs. Cycling conditions were: beginning with 94° C for 5 min, followed by 35 cycles of 94°C (1 min), 60°C (1 min), 72°C (1.5 min). The final extension at 72°C lasted again 5 min. The PCR products were electrophoresed in a 2% agarose gel, in a 0.5xTBE buffer. The PCR reaction yielded for all rice accessions examined a single fragment of about 250 bp in length, as could be detected after staining with ethidium bromide. The PCR products were purified with a QIA quick Gel extraction kit (Qiagen) and then directly sequenced, using BigDye terminator reactions (Applied Biosystems, Foster City, CA) that were run on an ABI 3700 automated sequencer. Reactions were performed by using the amplification primer. For each rice accession, forward and/or reverse sequencing reactions were run for sequence confirmation. DNA sequences of the wx gene, the presence of the wx microsatellite in the form of a (CT)n repeat in the exon 1 and the G/T polymorphism within a consensus donor site in intron 1, were visually observed on chromatograms. The two wx gene alleles, wxa and wxb have a GG and a GT sequence, respectively, at the splice donor site of intron 1. The two alleles were determined by using restriction enzyme cleavage. Five microliters of each PCR product were digested with Acc I (BRL) in a total volume of 20 μL at 37°C for 3 h. The restriction fragments were electrophoresed in TBE buffer using 3.5% agarose gel.
Amylose content determination: Seeds from a single plant of the samples that had been investigated for the wx microsatellite allele and for the wx gene allele were collected and used for amylose content determination, which has been done by a colorimetric assay of Juliano (1971).
RESULTS
wx microsatellite allele diversity in O. rufipogon: Seven wx microsatellite alleles, (CT)n repeats of n = 9, 10, 11, 12, 16, 17 and 18 at the wx locus, were detected in this collection of wild rice (Fig. 1), of which the alleles (CT) 10 and (CT)17 were the most frequent ones. The distribution of these alleles is shown in Table 1. The (CT)10 allele was not found in the population from northern Thailand (Chiang Rai Province), but was prominent in the population from the northeastern region. The (CT)17 allele was detected in all regions of the country and Lao PDR. The (CT)18 allele was found only in the north and northeast of Thailand. The (CT)12 and (CT)16 alleles were rare. They have only been detected in a strain of wild rice from a large swamp in central Thailand and a wild rice accession from a roadside ditch in the northeast of the country.
Wx microsatellite allele association with G/T polymorphism at the splice donor site of intron 1: To study the relationship between the wx microsatellite allele and the G/T polymorphism, all 212 rice strains were sequenced in the region containing the wx microsatellite and the splice donor site of intron 1. For G/T polymorphism the sequences were confirmed using the PCR-RFLP method, as described in Materials and Methods. It was found that six alleles (n = 9, 10, 11, 12, 16 and 17) showed the sequence AGGTATA in the splice donor site, while the allele (CT)18 had the sequence AGTTATA at this position. The allele (CT)17 had either AGGTATA or AGTTATA. The restriction enzyme Acc I cleaves the sequence AGGTATA. Thereby, the ca. 250 bp PCR product containing the wx microsatellite and the splice donor site of intron 1 is divided into two smaller fragments. This does not happen to the PCR products from the rice strains with the sequence AGTTATA in the splice donor site (Fig. 2). In a recent SNP analysis by Yamanaka et al. (2004) in wild rice (O. rufipogon) samples, collected from the Mekong Basin of the Indochina Peninsula in Southeast Asia, only the sequence AGGTATA, but not the sequence AGTTATA, was found.
Wx microsatellite allele association with amylose class in wild rice strains: Seeds of those rice strains carrying the eight genotypes, revealed by the linkage phenomenon between wx microsatellite allele and G/T polymorphism at the first intron, were used to determine their amylose content. It was found that the genotypes (CT)9/G, (CT)10/G, (CT)11/G, (CT)12/G, (CT)16/G, (CT)17/G, (CT)17/T and (CT)18/T showed variation of amylose contents (Table 2). The six genotypes (CT = 9G, 10G, 11G, 12G, 16G and 17G) were subjected to an Acc I digestion of the PCR products resulted in two small fragments. In contrast, the two rice strains carrying the allele (CT)17/T and the allele (CT)18/T, showed a low amylose content, with an average of 14.9 and 12.5%, respectively. In these two genotypes the enzyme could not digest the PCR products. Previously, polymorphism of wx microsatellite in the wx gene of rice can account for more than 82% of the variation reported in amylose content of non-glutinous rice (Ayres et al., 1997) and regarded as a molecular marker in determining amylose content. In addition, sixty non-glutinous cultivars were detected for variation of wx microsatellite alleles (Tan and Zhang, 2001). It was found that rice cultivars with high amylose content (>22%) carried alleles with low numbers of (CT)n repeats, n = 14; contrast to cultivars with low to intermediate amylose content carried alleles with high numbers of (CT)n repeats, n>=16.
 |
| Fig. 1: | The 200 bp fragments, amplified with primer waxy F and waxyR2, were sequenced and compared among eight haplotypes [(CT)n/G-T] of the rice waxy gene in wild rice O. rufipogon. The CT repeats and G/T polymorphism at the splice site of the intron 1 are indicated by underlined letters |
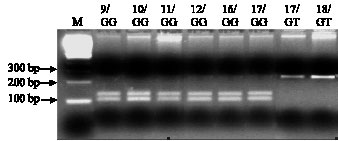 |
| Fig. 2: | Comparison between PCR-RFLP analysis and sequencing analysis for determining of G/T polymorphism in wild rice strains, lanes 2-9. Lane 1, M = 100 bp ladder |
| Table 2: | Waxy microsatellite allele at the Waxy gene and mean values of the amylose content (AC%) of O. rufipogon |
 | |
DISCUSSION
Previously, polymorphism of wx microsatellite in the wx gene of rice can account for more than 82% of the variation reported in amylose content of non-glutinous rice (Ayres et al., 1997) and regarded as a molecular marker in determining amylose content. In addition, sixty non-glutinous cultivars were detected for variation of wx microsatellite alleles (Tan and Zhang, 2001). It was found that rice cultivars with high amylose content (>22%) carried alleles with low numbers of (CT)n repeats, n<=14; contrast to cultivars with low to intermediate amylose content carried alleles with high numbers of (CT)n repeats, n>=16. In this study, a survey of 16 natural populations of wild rice O. rufipogon from Thailand and Lao PDR with respect to the wx microsatellite (CT)n at the wx locus showed seven wx microsatellite alleles with seven different repeat numbers: n = 9, 10, 11, 12, 16, 17 and 18. Wild rice strains which carried low numbers of (CT)n repeats, n<=16 had intermediate and high amylose content. These strains contained the AGGTATA sequence at the putative 5`-leader intron splice-junction site, except for the (CT)17 allele which was linked with either AGGTATA or AGTTATA sequence at the position. Wild rice strains carrying the genotype (CT)17/G showed high amylose content, whereas strains with low amylose content carried the genotype (CT)17/T and (CT)18/T. By summary, the finding of the present study is consistent with the previous reports. In addition, the association between wx microsatellite alleles and amylose content can be used for marker-assisted selection in breeding programs and in other studies. This might include studies on the origin of non-glutinous rice with low amylose content and glutinous rice cultivars.
The wild rice strains with the genotype (CT)17/T and (CT)18/T were found in populations from the Northern and Northeastern regions of Thailand. These two wx microsatellite alleles have been previously detected in glutinous and non-glutinous rice with low amylose content in Thailand (Prathepha and Baimai, 2004). These data suggest that rice cultivars containing the wx microsatellite alleles similar to those of wild rice might have been domesticated from populations of O. rufipogon. The alleles (CT)14 and (CT)20 were not found in the examined wild rice as reported by Bergman et al. (2001). Also, the four alleles (n = 8, 14, 19 and 20), present in the US rice germplasm, as reported by Ayres et al. (1997), were not detected in wild rice in this study. Furthermore, in the Thai rice cultivars, reported by Prathepha and Baimai (2004), five wx microsatellite alleles were detected (n = 11, 16, 17, 18 and 19). The (CT)19 allele was also not detected in the set of wild rice strains examined in this study. It is, therefore, the origin of cultivars with these alleles are in mystery, because the wild rice strains having these wx microsatellite alleles (CT = 8, 14, 19, 20) has not been identified in O. rufipogon to date. Specifically, the fact that these alleles were not observed in the sample of wild rice, suggesting either that the wild population that was ancestral to these alleles were not sampled in this collection, or that it is now extinct.
As the relatives of Asian cultivated rice and wild rice species (O. nivara, O. rufipogon) are commonly found and even coexist in rice field in tropical Asia and Ocenia (Vaughan, 1989). Although rice is primarily self-pollinated crop but cross-pollination does occur in rice. Natural hybridization between cultivated and wild rice were occurred as reported previously (Song et al., 2003b; Chen et al., 2004). Thus, rice-to-wild gene flow can have a significant impact on the wild populations. Based on the results of the present study, variation of a Simple Sequence Repeat (SSR) revealed by wx microsatellite of the waxy gene is a good candidate for use as an assisted DNA marker in studying gene flow from cultivated rice to wild rice or vice versa. This knowledge is the most importance both for genetic conservation of wild rice genetic resources and for risk assessment of ecological consequences of their hybrids. Due to O. rufipogon is considered not only to be the progenitor of O. sativa, but also it is the most important genetic resources for rice improvement in terms of its accessibility for gene transfer through sexual means (Oka, 1988).
Based on the present study, the wx microsatellite allele and the sequence variation at the splice junction of the rice waxy gene (i.e., haplotypes) of wild rice O. rufipogon are polymorphic enough to classify wild rice strains which have different amylose classes. It is therefore these DNA markers could be used as the marker-assisted selection for classifying grain amylose in wild rice.
ACKNOWLEDGMENTS
This research was supported by the TRF/BIOTEC Special Program for Biodiversity Research and Training grant BRT_R248003. The author is grateful to anonymous readers for critical reading manuscript and providing invaluable suggestions.
REFERENCES
- Akimoto, M., Y. Shimamoto and H. Morishima, 1999. The extinction of genetic resources of Asian wild rice, Oryza rufipogon Genet. Resour. Crop Evol., 46: 419-425.
CrossRef - Ayres, N.M., A.M. McClung, P.D. Larkin, H.F.J. Bligh, C.A. Jones and W.D. Park, 1997. Microsatellites and a single-nucleotide polymorphism differentiate apparentamylose classes in an extended pedigree of US rice germ plasm. Theor. Applied Genet., 94: 773-781.
CrossRefDirect Link - Bao, J.S., H. Coke and M. Sun, 2002. Microsatellites in starch-synthesizing genes in relation to starch physicochemical properties in waxy rice (Oryza sativa L.). Theor. Applied Genet., 105: 898-905.
Direct Link - Bergman, C.J., J.T. Delgado, A.M. McClung and R.G. Fjellstrom, 2001. An improved method for using a microsatellite in the rice waxy gene to determine amylose class. Cereal Chem., 78: 257-260.
CrossRef - Bligh, H.F.J., R.I. Till and C.A. Jones, 1995. A microsatellite sequence closely linked to the Waxy gene of Oryza sativa. Euphytica, 86: 83-85.
CrossRefDirect Link - Bligh, H.F., P.D. Larkin, P.S. Roach, C.A. Jones, H. Fu and W.D. Park, 1998. Use of alternate splice sites in granule-bound starch synthase mRNA from low-amylose rice varieties. Plant Mol. Biol., 38: 407-415.
CrossRefDirect Link - Cai, X.L., Z.Y. Wang, Y.Y. Xing, J.L. Zhang and M.M. Hong, 1998. Aberrant splicing of intron 1 leads to the heterogeneous 5' UTR and decreased expression of waxy gene in rice cultivars of intermediate amylose content. Plant J., 14: 459-465.
CrossRefDirect Link - Chen, L.J., D.S. Lee, Z.P. Song, H.S. Suh and B.R. Lu, 2004. Gene flow from cultivated rice (Oryza sativa) to its weedy and wild relatives. Ann. Bot., 93: 67-73.
Direct Link - Gao, L.Z., W. Chen, W.Z. Jiang, S. Ge, D. Hong and X. Wang, 2000. Genetic erosion in Northern marginal population of the common wild rice Oryza rufipogon griff and its conservation, revealed by the change of population genetic structure. Hereditas, 133: 47-53.
Direct Link - Gao, L.Z., B.A. Schaal, C.H. Zhang, J.Z. Jia and Y.S. Dong, 2002. Assessment of population genetic structure of common wild rice Oryza rufipogon griff detected by microsatellite DNA and allozyme loci. Theor. Applied Genet., 106: 173-180.
Direct Link - Gao, L.Z., 2004. Population structure and conservation genetics of wild rice Oryza rufipogon (Poaceae): A region-wide perspective from microsatellite variation. Mol. Ecol., 13: 1009-1024.
CrossRef - Hirano, H.Y., M. Eiguchi and Y. Sano, 1998. A single base change altered the regulation of the Waxy gene at the posttranscriptional level during the domestication of rice. Mol. Biol. Evol., 15: 978-987.
CrossRefDirect Link - Isshiki, M., K. Morino, M. Nakajima, R.J. Okagaki, S.R. Wessler, T. Izawa and K. Shimamoto, 1998. A naturally occurring functional allele of the rice waxy locus has a GT to TT mutation at the 5′splice site of the first intron. Plant J., 15: 133-138.
CrossRefDirect Link - Isshiki, M., Y. Yamamoto, H. Satoh and K. Shimamoto, 2001. Nonsense-mediate decay of mutant waxy mRNA in rice. Plant Physiol., 125: 1388-1395.
Direct Link - Prathepha, P. and V. Baimai, 2004. Variation of wx microsatellite allele, waxy allele distribution and differentiation of chloroplast DNA in a collection of Thai rice (Oryza sativa L.). Euphytica, 140: 231-237.
Direct Link - Song, Z.P., X. Xu, B. Wang, J.K. Chen and B.R. Lu, 2003. Genetic diversity in the Northernmost Oryza rufipogon populations estimated by SSR markers. Theor. Applied Genet., 107: 1492-1499.
CrossRef - Song, Z.P., B.R. Lu, Y.G. Zhu and J.K. Chen, 2003. Gene flow from cultivated roce to the wild species Oryza rufipogon under experimental field conditions. New Phytol., 157: 657-665.
Direct Link - Song, Z., B. Li, J. Chen and B.R. Lu, 2005. Genetic diversity and conservation of common wild rice (Oryza rufipogon) in China. Plant Species Biol., 20: 83-92.
CrossRef - Tan, Y.F. and Q.F. Zhang, 2001. Correlation of simple sequence repeat (SSR) variants in the leader sequence of the waxy gene with amylose content of the grain in rice. Acta Bot. Sin., 43: 146-150.
Direct Link - Yamanaka, S., I. Nakamura, K.N. Watanabe and Y.I. Sato, 2004. Identification of SNPs in the waxy gene among glutinous rice cultivars and their evolutionary significance during the domestication process of rice. Theor. Appl. Genet., 108: 1200-1204.
Direct Link - Zhou, H.F., Z.W. Xie and S. Ge, 2003. Microsatellite analysis of genetic diversity and population genetic structure of a wild rice (Oryza rufipogon Griff.) in China. Theor. Applied Genet., 107: 332-339.
Direct Link