
C. Bounphanousay
National Agriculture and Forestry Research Institute, Ministry of Agriculture and Forestry, P.O. Box 811, Vientiane, Lao PDR
P. Jaisil
Department of Plant Science and Agricultural Resources, Faculty of Agriculture, Khon Kaen University, Khon Kaen 40002, Thailand
K.L. McNally
T.T. Chang Genetic Resources Center, IRRI, DAPO Box 7777, Metro Manila 1301, The Philippines
J. Sanitchon
Department of Plant Science and Agricultural Resources, Faculty of Agriculture, Khon Kaen University, Khon Kaen 40002, Thailand
N.R. Sackville Hamilton
T.T. Chang Genetic Resources Center, IRRI, DAPO Box 7777, Metro Manila 1301, The Philippines
Asian Journal of Plant Sciences
Year: 2008 | Volume: 7 | Issue: 2 | Page No.: 140-148







ABSTRACT
The genetic diversity of 74 genotypes, including Black Glutinous Rice (BGR) from Lao`s germplasm was assessed using 24 microsatellite markers. A total of 75 alleles were detected at the 24 microsatellite markers. The number of alleles per marker varied from 2 to 7 with an average of 3.1 alleles per locus. The Gene Diversity (GD) and Polymorphism Information Content (PIC) ranged from 0.18 to 0.79 and 0.17-0.76, respectively and the Allele Frequency (AF) ranged from 0.36 to 0.90. The markers were able to classify rice genotypes into four groups; indica rices were put in the three groups while the other group consisted of tropical japonica rice. The first indica group (G1) included 24 genotypes of BGR and five genotypes of white rice. Most of genotypes in this group have thick culms, broad leaf blades, large and bold grain shapes and some of them have purple coloration on all vegetative parts. In other varieties with black pericarp, all other plant parts are green. Indica group 3 (G3) included 25 genotypes of BGR. The special characters of this subgroup were small and slender culms, narrow short leaves, purple leaf margins, purple leaf tips and purple stripes on leaf blades and sheathes. However, the markers used could not differentiate between LG 8215 and LG 7937. Indica group four (G4) consisted of eight genotypes of white rice and the four check varieties. Group (G2) consisted of five white rices, four BGRs and three check varieties.
PDF Abstract XML References Citation
How to cite this article
C. Bounphanousay, P. Jaisil, K.L. McNally, J. Sanitchon and N.R. Sackville Hamilton, 2008. Variation of Microsatellite Markers in a Collection of Lao`s Black Glutinous Rice (Oryza sativa L.). Asian Journal of Plant Sciences, 7: 140-148.
DOI: 10.3923/ajps.2008.140.148
URL: https://scialert.net/abstract/?doi=ajps.2008.140.148
DOI: 10.3923/ajps.2008.140.148
URL: https://scialert.net/abstract/?doi=ajps.2008.140.148
INTRODUCTION
Rice belongs to the genus Oryza in the grass family (Gramineae). There are 22 species in the genus, of which only two are cultivated: O. sativa, which was domesticated in the humid tropics of South and Southeast to East Asia and O. glaberrima, which was domesticated in the Niger basin in Africa (Khush, 1997). O. sativa was domesticated in south Asia at least 10,000 years ago (Zhang and Jiarong, 1998) and O. glabberima, was domesticated in West Africa between about 1500 and 800 BC (Murrey, 2004).
South and Southeast Asia is generally accepted as the center of origin and domestication of glutinous rice and glutinous rice varieties are grown in most countries in these areas that have long history of rice cultivation. Laos has the highest per capita production and consumption of glutinous rice in the world (Schiller et al., 2006). Lao PDR proved to have one of the richest biodiversities in rice and it appears to be the world`s biodiversity centre for glutinous (sticky) rice varieties (Anonymous, 2007). In Lao PDR, traditional varieties of black glutinous rice are named by farmers for a particular distinctive character or region and a single name may encompass a genetically diverse set of varieties. For example, rice varieties with coloured pericarps other than red are usually called Khao Dam or Khao Kam (Dark Rice or Black Rice) (Appa et al., 2002a).
The black rices of Laos are less in production and consumption, but they are not least in their importance. They have long been cultivated and are widely grown in small plots throughout the country for making special recipes in special occasions such as bamboo rice, rice cakes and special alcohol drinks. BGR varieties can command a premium price in both the international and domestic markets. Their superior characteristics, such as high eating quality and grain characteristics and good general adaptability to biotic and abiotic factors, make them popular among Lao farmers.
The Lao Ministry of Agriculture and Forestry (MAF) and the International Rice Research Institute (IRRI) had jointly collected 13, 992 samples of cultivated rice germplasm in Laos during 1995 to 2003 (Appa et al., 2002a, b) and 431 accessions (3.07% of the total collections) were classified as having coloured pericarps based on the information provided by the farmers at the time of collection of the germplasm and on the characterization data from the Agriculture Research Center (ARC), (ARC, 2003, 2004; Appa et al., 2003). This germplasm is currently conserved in the Lao`s national genebank and in the International Rice Germplasm Collection (IRGC).
In an attempt to best utilize this indigenous rice germplasm, several activities for short-term application have been initiated. These activities include primary characterization, evaluation in replicated yield trails (Inthapanya et al., 1995). For the longer term utilization of the germplasm, it is necessary to quantify genetic diversity among Lao BGR varieties and to classify the genetic differentiation among them at molecular level. Most studies have been conducted so far with non-glutinous rice, fragrant rice and white glutinous rice using several types of molecular markers. The lowest genetic diversity was observed among the traditional basmati varieties, whereas the evolved basmati varieties showed the highest genetic diversity by Simple Sequence Repeats (SSR) and Inter Simple Sequence Repeats (ISSR) assays (Nagaraju et al., 2002). Bao et al. (2006) reported that high levels of AFLP (78.3%) and ISSR (92.2%) polymorphism were found in 56 waxy (glutinous) rice accessions. In general, gene diversities in rice populations (landraces, old varieties, newly developed varieties, wild species and parental lines of hybrids) as measured by different types of molecular markers ranged from 32.0 to 70.0 depending on methods used and rice populations under study (Giarrocco et al., 2007; Pessoa et al., 2007; Xu et al., 2004; Yu et al., 2003; Coburn et al., 2002; Ni et al., 2002) except for very low genetic diversity (PIC = 0.001) of landrace accessions in high altitude regions of Nepal (Bajracharya et al., 2006).
Most studies identified indica-japonica pattern of genetic diversity of Asian rice. In a study on a study of a wide range of rice genotypes using nuclear SSRs and two chloroplast loci, Garris et al. (2005) found five distinct groups corresponding to indica, aus, aromatic, temperate japonica and tropical japonica rices. They also found that indica was closely related with aus, whereas tropical japonica, temperate japonica and aromatic rices were closely related. In a study in Indonesia conducted on traditional and improved varieties, Thomson et al. (2007) found that indica rice (68%) predominated over tropical japonica rice (32%) in total number of accessions studied but gene diversities were rather similar (0.53 and 0.56 for indica and tropical japonica, respectively). Using combined AFLP and SSR markers, Bao et al. (2006) could clearly differentiate indica and japonica groups of waxy rice accessions.
Unfortunately, this very important information for black glutinous rice is not available in the literature. This is because most rice breeding programs have been focused on non-glutinous rice and breeding of glutinous rice especially for black glutinous rice is far behind. The objective of this study was to use SSR markers to quantify genetic diversity among Lao BGR varieties and classify the genetic differentiation among them. The information will be useful for future breeding program, conservation and effective management of black rice genetic resources.
MATERIALS AND METHODS
Plant materials: The research was undertaken in 2006 at the IRGC at the International Rice Research Institute (IRRI), Philippines. Rice varieties used for this study consisted of 56 varieties of Oryza sativa provided by the Lao`s national genebank (Table 1) and 18 varieties of standard white rice supplied by IRGC. Fifty three varieties out of the total of 74 varieties were tested and classified into black rice. The remaining consisted of tropical japonica, indica rice and white glutinous and non-glutinous rice.
DNA extraction: Total genomic DNA was extracted from young leaves using a modification of the method described by Fulton et al. (1995). Approximately 5 mg of young leaf tissue was frozen in liquid nitrogen, followed by grinding using a plastic bit attached to an electric drill before addition of extraction buffer, which was different from that of Fulton et al. (1995) who used mortar and pestle. Other steps in extraction followed Fulton et al. (1995). After extraction, pellets were dissolved in 150 μL TE (10 mM Tris, 1 mM EDTA, pH 8). DNAs were quantified by gel densitometry on agarose gels using Lambda DNA as a standard followed by normalization to concentration of 5 ng μL-1 prior to use in PCR.
PCR amplification: Polymerase chain reactions using microsatellite primers were carried out in a final volume of 10 μL. The PCR reaction mixture contained 1 μL 10xTBE buffer (containing 100 mM Tris-HCl pH 8.5, 50 mM KCl), 1 μL 15 mM MgCl2, 1 μL of 5 mM dNTP mixture, 1 μL each of 10 mM forward and reverse primers, 1 unit (0.5 μL) of Taq DNA polymerase, 10 ng of DNA template and ddH2O to reach the final reaction volume of 10 μL. Reactions were overlaid with mineral oil.
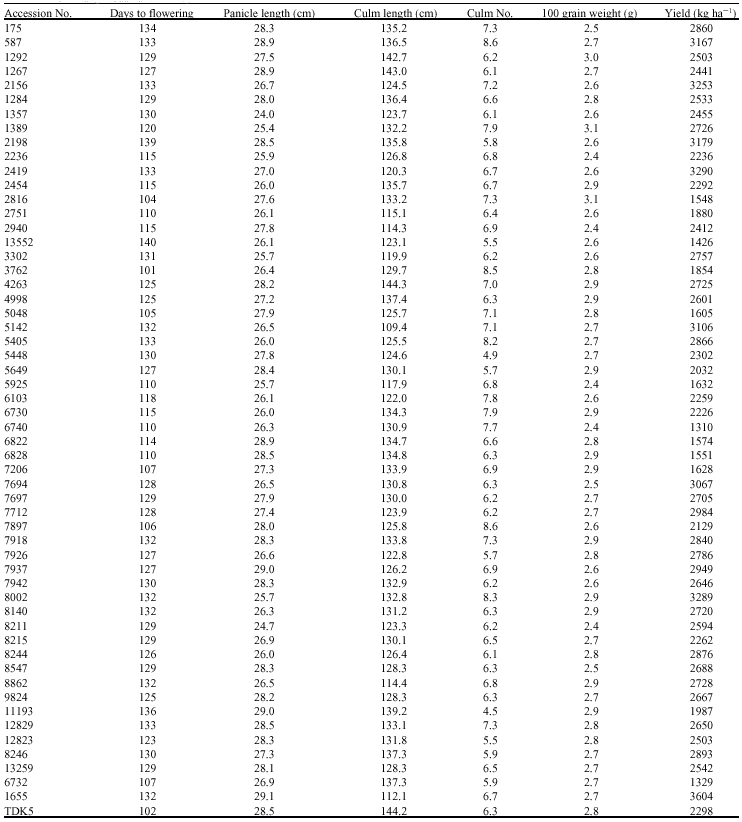
| Table 1: | Some agronomic characters of 53 accessions of lowland black glutinous rice and three varieties of Lao white rice (6732 = Kai Noi Leuang, 1655 = Hom Nang Nouan and TDK5) |
 | |
To optimize the PCR amplification conditions, experiment were carried out with varying quantity of MgCl2 (15 mM) and Taq DNA polymerase, two different quantity of MgCl2 and Tag polymerise were used (1-1.5 and 0.5-1 μL) respectively.
The 35 SSR markers were screened in this study. Eleven SSR markers were not polymorphic and discarded. Twenty four polymorphic markers were used to amplify DNA products (Table 2). These markers were reported previously (Temnykh et al., 2000). Amplifications were carried out using MJR DNA Engine Dyad® thermal cyclers. The PCR programs consisted of the following steps: initial denaturation for 5 min at 94°C, followed by 40 cycles of 1 min denaturation at 94°C, 1 min annealing at 55°C, 2 min extension at 72°C, followed by a final extension of 5 min at 72°C and holding at 15°C until recovery. For some specific microsatellites, a different annealing temperature of 53, 57, 61 or 63°C was used as indicated in Table 2.
| Table 2: | Total number of alleles, allele length in bp, gene frequency and gene diversity of 24 SSR markers assayed in 74 rice genotypes |
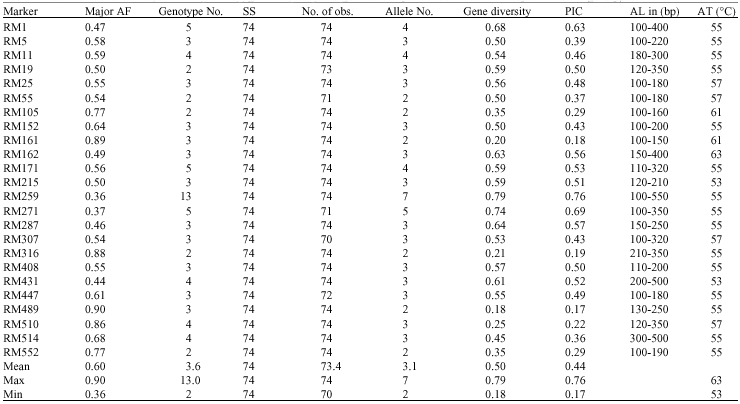 | |
| Remark: AF = Allele Frequency, AL = Allele Length in bp, SS = Sample Size, AT = Annealing Temperature and PIC = Polymorphic Information Content | |
Amplification products were separated by non-denaturing PAGE on mini vertical units using either 6.5 or 8% polyacrylamide gels with 1x TBE buffer at 100 volts for 1.5 h. After electrophoresis, gels were stained with ethidium bromide and visualized by UV illumination. Gel images were photographed using Bio-Rad GelDoc system. The molecular weights of PCR products were estimated relative to a 1 kb ladder that served as the size standard.
Data analysis: Summary statistics (PIC, GD and AF) on the marker data were calculated using PowerMarker 3.25 (Liu and Muse, 2005).
The allelic state encoded SSR data were used to calculate a matrix of genetic distances between all pairs of varieties using simple matching coefficient under DARwin® 4.0 Release (2007) (Perrier et al., 2003). Cluster analysis was accomplished on this matrix by the unweighted neighbor joining option.
RESULTS
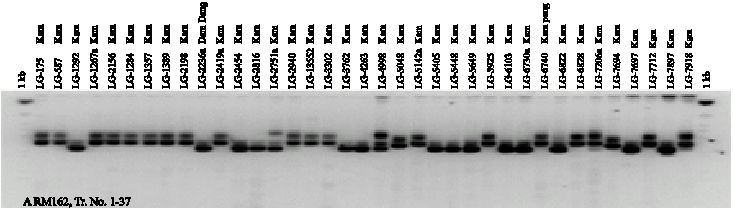
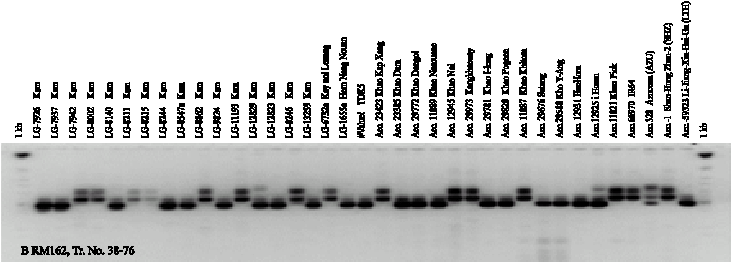
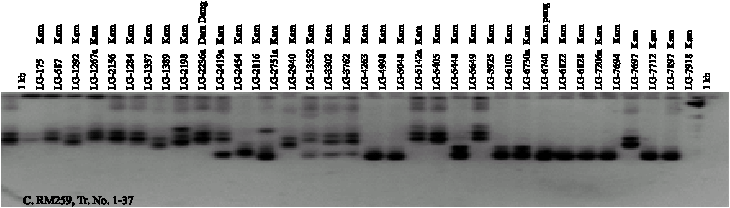
Twenty-four SSR markers were used for amplifying DNA segments from genomic DNA of 74 genotypes. A total of 75 alleles were scored from the 24 SSR markers. The number of alleles per primer ranged from 2 to 7. On average, 3.1 alleles per locus were observed. The Gene Diversity (GD) ranged from 0.18-0.79. The Polymorphism Information Content (PIC) ranged from 0.17 to 0.76 and the Allele Frequencies (AF) ranged from 0.36 to 0.90 (Table 2). RM259 SSR was the highest polymorphic marker with 7 alleles. The size of the amplified segment ranged from 100-150 bp for RM161 to 100-550 bp for RM259. Number of genotypes from 24 SSR markers ranged from 2-13. The PCR products amplified by RM259, RM307 and RM162 are shown in Fig. 1. The number of alleles detected by RM171, RM271 and RM259 was slightly higher than the average with 4-7 alleles.
The microsatellite markers were able to distinguish between different rice genotypes. The high degree of polymorphism of microsatellite markers allows rapid and efficient identification of rice genotypes. The microsatellite markers classified the rice genotypes into four groups; indica accessions were put in the three groups while the other group consisted of tropical japonica rice. Genotypes in indica group one (G1) included 24 genotypes of BGR and five genotypes of white rice. Most of genotypes in this group have thick culms, broad leaf blades, large and bold grain shapes and some of them have purple coloration on all plant parts. In other varieties with black pericarp, other plant parts including glumes and leaves are green color. Indica group 3 (G3) included 25 genotypes of BGR. The special characters of this subgroup were small and slender culms, narrow short leaves, purple leaf margins, purple leaf tips and purple stripes on leaf blades and sheaths. However two genotypes were not distinguished from each other (LG 8215 and LG 7937), these might be of the same genotype. Indica group four (G4) consisted of eight genotypes of white rice, which were green color with tall plant parts and included the four check varieties (Hom Nang Nouan, TDK5, IR64 and Shan-Huang Zhan-2). Tropical japonica group (G2) consisted of 5 genotypes of Lao white rice varieties and 4 genotypes of BGR and three check varieties (Khao Kai Noi Leuang, Azucena and Li-Jiang-Xin-Hei-Gu). Hence, the four BGR genotypes (LG6730, LG7260, LG8140 and LG13259) in this group should be tropical japonica types (Fig. 2).
 | |
 | |
 | |
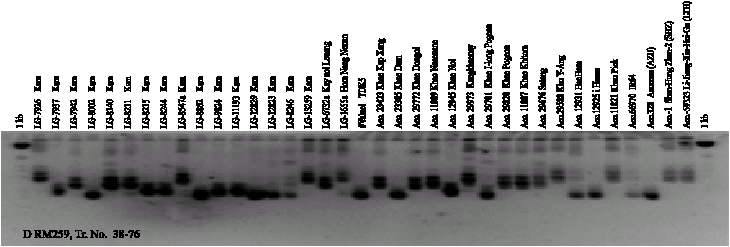 | |
| Fig. 1: | SSR banding patterns of 53 black glutinous rice and 21 white rice generated from RM162 (A and B) and RM259 (C and D) |
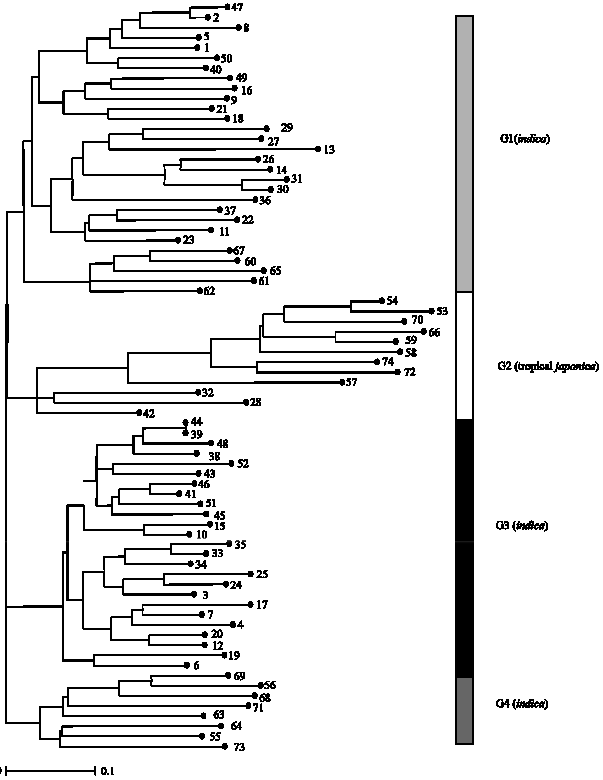 | |
| Fig. 2: | Dendrogram of 74 rice genotypes, constructed using UPGMA based on simple matching coefficient using allelic states |
DISCUSSION
Glutinous rice is a staple food for Lao people and black glutinous rice is also important as food for special occasions and special recipes. Black glutinous rice also serves as a valuable genetic resource for genes with economic importance such as resistance to abiotic and biotic stress, table quality and starch quality for a variety of value-added products. As a part of the center of origin and domestication of glutinous rice, high genetic diversity of this type of rice would be expected in the samples of total accessions collected in different regions of the country. The main objective of this research was to understand genetic variability of the indigenous black rice germplasm. The other objectives were to select parental genotypes for potential use in future breeding program.
Out of 35, 24 SSR primers revealed rather moderate genetic diversity of black glutinous rice as indicated by relatively low alleles per locus (3.1), GD (0.50) and PIC (0.44), when compared with the results of those available in the literature. By using 26 SSR markers, Giarrocco et al. (2007) found high alleles per locus (8.4) low pair-wise genetic similarity (0.32) and high PIC (0.69) in 69 rice cultivars with historical importance in Argentina. By using 47 RAPD markers, Singh et al. (2003) obtained a total number of 275 alleles (75 allels in present study) in 21 rainfed lowland rice genotypes consisting of historic and modern cultivars. In general, genetic diversities reported in previous studies (Xu et al., 2004; Bao et al., 2006; Pessoa et al., 2007; Thomson et al., 2007) were higher than this study but the results cannot be compared directly because neither of the previous research was conducted with black glutinous rice. However, Bajracharya et al. (2006) found low diversity in landrace population in Nepal with diversity index of 0.23 and these genotypes might derive from single origin. The lower genetic diversity might be caused by the following reasons. First, the samples represented only lowland black glutinous rice excluding upland black glutinous rice. Second, the samples are not well represented the total genetic variability because of low accessibility to localized areas and, if this is the case, resampling in remote sites is recommended to collect additional samples. Third, there are differences in rice populations and methods used in many studies.
Out of 35, 11 SSR primers failed to differentiate black glutinous rice samples indicated that genetic variability was not high and a large number of randomly distributed SSR primers are required to identify the genotypes. Ni et al. (2002) suggested that 30 SSR primers were needed to stabilize the dendrogram of 38 rice accessions. The other possible reasons for low genetic diversity in black glutinous rice are that preferential selection of farmers and genetic erosion by introduction of modern varieties might also be the causes. This highlights the importance of genetic conservation of black glutinous rice germplasm for future use. From the preliminary study on agronomic characters (Table 1), the accessions of BGR had high variations in many characters such as yield, days to flowering, panicle length and culm length. Improvement of economically important characters of this germplasm can be achieved but might be in limited extent because of low genetic diversity. Introduction of genetic materials from diverse sources is required.
Typical indica-japonica grouping pattern was clearly identified by the SSR dendrogram. Three groups formed an indica main group, whereas only one group represented japonica type. Higher diversity was found in idica rice than in tropical japonica rice. The results corresponded to the widely accepted grouping pattern of Asian rice (Giarrocco et al., 2007; Yu et al., 2003; Ni et al., 2002; Glaszmann, 1987) in which six groups constitute O. sativa species (Glaszmann, 1987). The results were in agreement with the study of Prathepha and Baimai (2004) who found that almost all of glutinous upland rice genotypes grown by various ethnic groups in northern Thailand were characterized as japonica type, whereas most of rainfed glutinous lowland varieties from other regions of Thailand were indica. A study with Thai and Lao black glutinous rice found that based on two growing conditions, rainfed lowland and rainfed upland, chloroplast DNA type was distinct from each other (Prathepha, 2007).
To better utilize black glutinous rice in the country and achieve breeding goal, genetic diversity of upland glutinous rice should be studied and compared with this study. Black glutinous rice in other countries should be studied and introduced to broaden germplasm base of black glutinous rice.
ACKNOWLEDGMENTS
We thank Mr. Phumi INTHAPANYA Director of Agricultural Research Center of Lao PDR, for seed samples of Lao accessions. Financial support from IRRI`s Training Center is gratefully acknowledged.
REFERENCES
- Appa, R.S., C. Bounphanousay, J.M. Schiller, A.P. Alcantara and M.T. Jackson, 2002. Naming of traditional rice varieties by farmers in the Lao PDR. Genet. Res. Crop Evol., 49: 83-88.
CrossRef - Appa, R.S., C. Bounphanousay, J.M. Schiller and M.T. Jackson, 2002. Collection, classification and conservation of cultivated and wild rices of the Lao PDR. Genet. Res. Crop Evol., 49: 75-81.
CrossRef - Bao, J., C. Harold and S. Mei, 2006. Analysis of genetic diversity and relationships in waxy rice (Oryza sativa L.) using AFLP and ISSR Markers. Genet. Res. Crop Evol., 53: 323-330.
Direct Link - Bajracharya, J., K.A. Steele, D.I. Jarvis, B.R. Sthapit and J.R. Witcombe, 2006. Rice landrace diversity in Nepal: Variability of agro-morphological traits and SSR markers in landraces from a high-altitude site. Field Crops Res., 95: 327-335.
Direct Link - Coburn, J.R., S.V. Temnykh, E.M. Paul and S.R. McCouch, 2002. Design and application of microsatellite marker panels for semiautomated genotyping of rice (Oryza sativa L.). Crop Sci., 42: 2092-2099.
Direct Link - Fulton, T.M., J. Chunwongse and S.D. Tanksley, 1995. Microprep protocol for extraction of DNA from tomato and other herbaceous plants. Plant Mol. Biol. Rep., 13: 207-209.
CrossRefDirect Link - Garris, A.J., T.H. Tai, J. Coburn, S. Kresovich and S. McCouch, 2005. Genetic structure and diversity in Oryza sativa L. Genetics, 169: 1631-1638.
CrossRefPubMedDirect Link - Giarrocco, L.E., M.A. Marassi and G.L. Salerno, 2007. Assessment of the genetic diversity in Argentine rice cultivars with SSR markers. Crop Sci., 47: 851-858.
Direct Link - Glaszmann, J.C., 1987. Isozymes and classification of Asian rice varieties. Theoret. Appl. Genet., 74: 21-30.
CrossRefDirect Link - Inthapanya, P., J.M. Schiller, S. Sarkarung, T. Kupkanchanakul and V. Phannourath, 1995. Varietal improvement strategies for the rainfed lowland environment of the Lao PDR: 1995-2000: Fragile lives in fragile ecosystems. Proceedings of the 1995 International Rice Research Conference, February 13-17, 1995, IRRI, Manila, pp: 767-782.
- Khush, G.S., 1997. Origin, dispersal, cultivation and variation of rice. Plant Mol. Biol., 35: 25-34.
CrossRef - Liu, K. and S.V. Muse, 2005. PowerMarker: An integrated analysis environment for genetic marker analysis. Bioinformatics, 21: 2128-2129.
CrossRefDirect Link - Murray, S.S., 2004. Searching for the origins of African rice domestication. Antiquity, Vol. 78, No. 300.
Direct Link - Nagaraju, J., M. Kathirvel, R.R. Kumar, E.A. Siddiq and S.E. Hasnain, 2002. Genetic analysis of traditional and evolved Basmati and non-basmati rice varieties by using fluorescence-based ISSR-PCR and SSR markers. Proc. Natl. Acad. Sci., 99: 5836-5841.
CrossRefDirect Link - Ni, J., P.M. Colowit and D.J. Mackill, 2002. Evaluation of genetic diversity in rice subspecies using microsatellite markers. Crop Sci., 42: 601-607.
Direct Link - Pessoa-Filho, M., A. Belo, A.A. Alcochete, P.H. Rangel and M.E. Ferreira, 2007. A set of multiplex panels of microsatellite markers for rapid molecular characterization of rice accessions. BMC Plant Biol., Vol. 7.
CrossRefDirect Link - Prathepha, P. and V. Baimai, 2004. Variation of wx microsatellite allele, waxy allele distribution and differentiation of chloroplast DNA in a collection of Thai rice (Oryza sativa L.). Euphytica, 140: 231-237.
Direct Link - Preecha, P., 2007. An assessment of Wx microsatellite allele, alkali degradation and differentiation of chloroplast DNA in traditional black rice (Oryza sativa L.) from Thailand and Lao PDR. Pak. J. Biol. Sci., 10: 261-266.
CrossRefPubMedDirect Link - Singh, S., S. Sarkarung, R.K. Singh, O.N. Singh, A.K. Singh, V.P. Singh, H.S. Bhandari, W. Xu and Z. Li, 2003. Genetic diversity in rainfed lowland rice genotypes as detected by RAPD primer: Advances in Rice Genetics. Proceedings of the 4th International Rice Genetics Symposium, October 22-27, 2003, IRRI, Los Banos, pp: 200-203.
- Temnykh, S., W.D. Park, N. Ayres, S. Cartinhour and N. Hauck et al., 2000. Mapping and genome organization of microsatellite sequences in rice (Oryza sativa L.). Theor. Applied Genet., 100: 697-712.
CrossRefDirect Link - Thomson, M.J., E.M. Septiningsih, F. Suwardjo, T.J. Santoso, T.S. Silitonga and S.R. McCouch, 2007. Genetic diversity analysis of traditional and improved Indonesian rice (Oryza sativa L.) germplasm using microsatellite markers. Theor. Applied Genet., 114: 559-568.
Direct Link - Xu, Y., H. Beachell and S.R. McCouch, 2004. A marker-based approach to broadening the genetic base of rice in the USA. Crop Sci., 44: 1947-1959.
CrossRefDirect Link - Yu, S.B., W.J. Xu, C.H.M. Vijayakumar, J. Ali and B.Y. Fu et al., 2003. Molecular diversity and multilocus organization of the parental lines used in the international rice molecular breeding program. Theor. Applied Genet., 108: 131-140.
Direct Link - Zhang, W. and J. Yuan, 1998. A preliminary study of ancient excavated rice from Yuchanyan site, Dao County, Hunan province, PR China. Acta Agron. Sin., 24: 416-420.
Direct Link