
Davarynejad Gholam Hossein
Department of Horticultural Science, College of Agriculture, Ferdowsi University, P.O. Box 91775-1163, Mashhad, Iran
Shahriari Farajollah
Department of Plant Biotechnology and Breeding, Ferdowsi University, P.O. Box 91775-1163, Mashhad, Iran
Hamid Hassanpour
Department of Horticultural Science, College of Agriculture, Ferdowsi University, P.O. Box 91775-1163, Mashhad, Iran
Asian Journal of Plant Sciences
Year: 2008 | Volume: 7 | Issue: 1 | Page No.: 109-112







ABSTRACT
This research was initiated to survey the peroxidase isozyme profiles of 17 pear (Pyrus communis L.) cultivars and compare their peroxidase profiles to Quince A (QA) (Cydonia oblonga Mill.) rootstock. Additionally, samples from the graft union of pear cultivars and quince rootstocks were analyzed for peroxidase isozymes to identify possible association with compatible graft combinations. The total amount of starch content was also considered as another marker to identify the scion-rootstock compatibility. Two anodal peroxidase A (RF = 0.86), present in Beurre Hardy and QA and B (RF = 0.68), present in Beurre Hardy and absent in QA was considered as rootstock and scion relationship. The presence of A band in the graft union was an indication of compatibility of Beurre Hardy and Passa Crassana pear cultivars with QA, whereas, the absence of this band was related to incompatibility of Dargazi, Shahmivah and Torsh. Although the B band was present in Beurre Hardy, the absence of this band in QA was highly related to incompatibility of Shekari, Lizbon, Khoje asiabrak, Domkaj, Tabrizi, Koshia and Bolghari pear cultivars. The amount of starch accumulation near grafting sites (above and below) showed that Natanz, Dargazi, Shahmivah, Aloret and Torsh pear cultivars compared to others had greater amount of starch above the grafting sites.
PDF Abstract XML References Citation
How to cite this article
Davarynejad Gholam Hossein, Shahriari Farajollah and Hamid Hassanpour, 2008. Identification of Graft Incompatibility of Pear Cultivars on Quince Rootstock by Using Isozymes Banding Pattern and Starch. Asian Journal of Plant Sciences, 7: 109-112.
DOI: 10.3923/ajps.2008.109.112
URL: https://scialert.net/abstract/?doi=ajps.2008.109.112
DOI: 10.3923/ajps.2008.109.112
URL: https://scialert.net/abstract/?doi=ajps.2008.109.112
INTRODUCTION
Graft incompatibility in fruit trees is one of the greatest obstacles in rootstock breeding. This incompatibility could be the result of genetical, physiological or anatomical aspects (Hartmann et al., 1997). Graft incompatibility might be due to the absence of differentiation of callus tissues into new phloem tissues or necrosis of the cells in the site of scion as reported by Moore (1983). These phenomena can cause a miss-joining between rootstock and scion, leading to lack of lignifications of cells in the site of scion. Some incompatibilities have delayed effects, as observed to the black-line condition of English walnut grafted on Black walnut, appeared only after twenty or more years of satisfactory performance. Most incompatibilities of orchards trees, however, were apparent at an early age (Westwood et al., 1971). In the former decade, the swelling in the site of scion, yellowish of leaflet, reduction of vegetative growth and differences in growth rate between rootstock and scion was known as graft incompatibility (Hartmann et al., 1997). These markers have some disadvantages because it could take several years for the appearance of these symptoms and sometimes there is no correlation between anatomical observations and graft incompatibility (Andrews and Marquez, 1993). Analysis of Isozymes between rootstock and scion can be used for prediction of rootstock incompatibility as reported by Santamour et al. (1986). Graft compatibility occurs when the isozyme band pattern between scion and rootstock is similar. In this situation the vessel linears are quite desirable. It has been also suggested by Gulen et al. (2002), that one anodal Peroxide A can be associated with compatible graft combination. Moreover, they concluded that presence of isoperoxidases A and B in the graft union tissues may be used as an indicator to predict a compatible graft between pear and quince rootstock. On the other hand, starch accumulation above the scion or sites shortage under this site might cause decay of phloem as shown by Mosse (1962). Graft components with poor growth, not only leads to structural abnormalities in the site of scion, but also is correlated with irregularities in starch distribution.
Formation of assimilates and their mobility between root and shoot is highly affected by the level of rootstock and scion compatibility. The ratio of starch substances to total dry matter in leaves and shoots is linearly reduced by severity of incompatibility. This phenomenon is a good marker for evaluation of rootstock incompatibility as indicated by Herrero (1951). Starch distribution between different parts of grafting trees has shown that there is a relationship between incompatibility and starch metabolism. It was observed by Mendel and Cohen (1967), that in spite of incompatibility symptoms between grafted Cherry on Mahleb rootstock, the amount of starch above and below the grafting site was the same.
In Iran, quince and pear seedlings are common rootstocks for pear cultivars. According to the morphological studies in commercial orchards, it was found by Davarynejad and Davarynejad (2007) that the most important Iranian pear cultivars such as: Natanz, Shekari and Sebri are quite incompatible with QA. This phenomenon is normally occurring very late (8 to 10 years old or even more). It caused root out and replanting of some commercial pear orchards. This study was carried out to predict the compatibility of pear cultivars with quince in early stage of growth, using Isozyme markers as well as starch accumulation above and below the site of grafting in the plant tissue as an another indication of incompatibility
MATERIALS AND METHODS
Plant material: Pot experiments were conducted at greenhouse throughout the year of 2005. Rootstocks were budded after 3 months. The pear cultivars used in this experiment were: Beurre Hardy, Bolghari, Dargazi, Domkaj, Felestini, Jifard, Khoje Asiabrak, Koshia, Lizbon, Natanz, Passa Crassana, Shahmivah, Shekari, Torsh, Tabrizi and Spadana. Beurre Hardy cultivar was used as control.
Isozyme studies: Samples from a thin layer of cambium tissue of rootstock, scion and graft union were taken at July 2006 for isozyme analysis. Samples were first frozen using liquid nitrogen and then kept at -80 °C.
Extraction was performed on peroxides enzyme using Gulen et al. (2002) methodology. A sample of grinded tissue (60 mg) was initially mixed with extraction buffer (100 mM potassium phosphate, 30 mM boric acid; 50 mM L-ascorbic acid; 17 mM sodium metabisulfite; 16 mM dithiocarbamic acid; 1 mM EDTA (ethylenediaminetetraacetic acid) and 4% (w/v) PVP-40 (polyvinylpyrrolidone) and final pH was readjusted to 7.5 with NaOH) and then vortexed for few minutes and finally centrifuged at 16000 g for 30 min at 4 °C. At the end of centrifugation, the supernatant was used for electrophoresis.
Polyacrylamide gel electrophoresis (PAGE): Electrophoresis was performed with some modification of the method of Gulen et al. (2002). In this system, the upper and lower gel was used with concentrations of 5% and 12.5%, respectively. Electrophoresis was performed at 20 mA until the samples entered the separating gel (about 20 min) and then rerun with the same electrical current for about 20-40 min.
Gel staining: Gels were stained for peroxides using the method described by Wendel and Weeden (1989). Gels were then rinsed with distilled water, fixed and stored in 10% glycerol. The relative distance (RF value) of the bands on the gel was calculated as described by Manganaris and Alston (1992), using RF = 1.0, the distance to the fastest band (or the finished point of the running) and RF = 0.0, the starting point of the running
Starch estimation: Starch was extracted according to the method of Zapata et al. (2004) using tissue bark and wood 1 cm above and below grafting sites. Samples of tissue were first grinded using liquid nitrogen and then mixed with di-methyl sulfoxidase solution and left to stand for 1 h at 100 °C.
After centrifuging for 15 min at 12000 g, the amount of starch was measured using spectrophotometer at 620 nm wave length. For reading the light absorbent, extracts first reacted with iodides solution and were placed in the spectrophotometer. This experiment was conducted as completely randomize design with 3 replications per treatments. Duncan mean comparison was done using DNMRT.
RESULTS AND DISCUSSION
Native PAGE profiles of isoperoxidases of non budded scion and rootstock revealed predominantly anodal isoperoxidases (Table 1). Analysis of profiles revealed on isoperoxidases band (band A RF = 0.88) that was present in Beurre Hardy (a compatible pear cultivar). Another isoperoxidase band (band B RF = 0.68) was also observed in Beurre Hardy scion, so this cultivar can be used for comparing to other cultivars. The result of this experiment showed that the band B is absent in QA, so band A was the common band in Beurre Hardy scion and QA rootstock.
Relationship between these two bands and compatibility of pear cultivars on QA has also been confirmed by Gulen et al. (2002, 2005a, b), Harkin and Obst (1973), Hartmann et al. (1997) and Herrero (1951).
In this experiment, there was no evidence showing that B band in scion cultivar is related to incompatibility,
| Table 1: | Isozyme bands peroxidase in QA and pear cultivars with reaction of grafted rootstock |
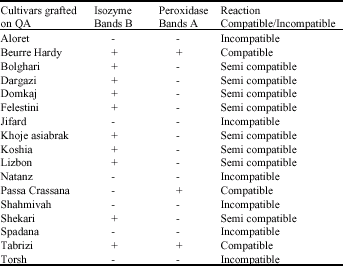 | |
it can be interpreted that the presence of B band is related to semi-compatibility of pear cultivars on QA (Table 1). Ermel et al. (1999) reported that Passa Crassana cultivar which is quite compatible with QA, include band A, but lacks band B. The bands of this cultivar relatively were similar to bands of QA, indicating that this cultivar was compatible with QA.
Gulen et al. (2002) reported that graft compatibility can occur only in the presence of A band. Presence or absence of peroxidases was basically due to the modification in gene expression. These changes probably occur by a signal, produced in the contact area of rootstock and scion.
Morphological studies in commercial orchards by Davarynejad and Davarynejad (2007) revealed that Natanz cultivar is quite incompatible with QA. Isozyme studies on this cultivar showed that, A and B bands were not present in this cultivar (Table 1). The results of morphological and starch content can be confirmed by the results of Isozyme studies. The specific role of peroxidase has not been clearly understood.
According to the morphological studies, Torsh Pear cultivar was known as incompatible cultivar. The isozyme study was also shown that none of the two bands A and B was present in this cultivar (Table 1). The absence of B band in this cultivar showed that this band could be involved in graft incompatibility, so the presence of this band can be used as a marker of compatibility.
Starch content and graft incompatibility is shown in Table 2. The highest amount of starch accumulation above graft site belonged to Aloret cultivar. Morphological studies conducted by Davarynejad and Davarynejad (2007) confirmed that Shahmivah and Dargazi were two incompatible cultivars with QA. These cultivars showed the higher starch accumulation above
| Table 2: | Amount of starch in bark, below and above graft union in different combination of pear cultivars on quince A (on the base of fresh weight) |
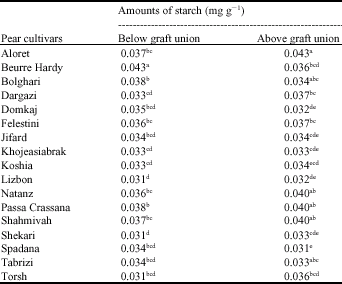 | |
| Duncan`s New Multiple Rang Test (DNMRT) was used for mean differences between cultivars for starch accumulation. Different letter(s) indicate significant differences between treatments (α = 0.01), CV = 13.5, N = 3 | |
the graft union. Torsh pear cultivar also showed the higher amount of starch accumulation above the grafted area (Table 2).
The higher starch accumulation in Natanz cultivar can be directly attributed to incompatibility of this cultivar with QA. Moing and Gaudillere (1992) also reported that, seventy eight days after grafting, concentration of sorbitol in the roots of incompatible graft, was lower than the compatible one, whereas, soluble sugars and starch were accumulated in the incompatible graft. This issue explains that the starch accumulation above the graft area could be an indication of graft incompatibility. Rem and Rabert (1987) suggested that starch accumulation above the graft site and lack of starch under this site may cause phloem decay and leads to vessel closure. The lack of starch below the graft site is certainly rare.
Table 2 shows that the highest amount of starch below graft union was observed in Beurre Hardy and the lowest amount was belonged to Shekari, Lizbon and Torsh cultivars. The highest amount of starch above the grafting site belonged to Aloret and the lowest amount observed in Spadana cultivars.
This study indicated that Natanz, Dargazi, Shahmivah, Aloret and Torsh pear cultivars accumulated more starch above the site of graft union, whereas, starch accumulation for other cultivars was absent or trace. It is noticeable that a cultivar may show symptoms of incompatibility without any starch accumulation above the grafting site, as suggested by Rem and Rabert (1987). However, in their study, when cherry was grafted on Mahleb, the symptoms of incompatibility was noticed, but no difference was observed between starch content above and below the site of mechanical treatments. This event shows that starch accumulation may not occur above the graft union in some cultivars, but the symptoms of incompatibility can be easily seen.
In conclusion, this study revealed that Natanz, Dargazi, Shahmivah, Aloret and Torsh pear cultivars were completely incompatible with QA, whereas Beurre Hardy and Passa Crassana were quite compatible. Establishing of a commercial and extensive orchard of such cultivars (Natanz, Dargazi and Shahmivah), requires to propagate them by compatible interstem or by their own roots.
REFERENCES
- Ermel, F.F., J. Kervella, A.M. Catesson and J.L. Poessel, 1999. Localized graft incompatibility in pear/quince (Pyrus communis/Cydonia oblonga) combinations: multivariate analysis of histological data from 5-month-old grafts. Tree Physiol., 19: 645-654.
PubMed - Gulen, H., R. Arora, A. Kuden, S. Krepbs and J. Postman, 2002. Peroxidase isozyme profiles in compatible and incompatible pear-quince graft combinations. J. Am. Soc. Hortic. Sci., 127: 152-157.
Direct Link - Gulen, H., M. Celik and A. Eris, 2005. Cambial isoperoxidases related to graft compatibility in pear/quince graft combinations. Turk. J. Agric., 29: 83-89.
Direct Link - Gulen, H., A. Kuden, J. Postman and R. Arora, 2005. Total protein content and SDS-PAGE in pear scions grafted on QA and pear seedling rootstocks. Turk. J. Agric., 29: 91-96.
Direct Link - Harkin, J.M. and J.R. Obst, 1973. Lignification in trees: indication of exclusive peroxidase participation. Science, 180: 296-298.
CrossRefDirect Link - Herrero, J., 1951. Studies of compatible and incompatible graft combinations with special reference to hardy fruit trees. J. Hortic. Sci., 26: 186-237.
CrossRefDirect Link - Manganaris, A.G. and F.H. Alston, 1992. Inheritance and linkage relationships of peroxidase isoenzymes in apple. Theor. Applied Genet., 83: 392-399.
CrossRef - Mendel, K. and A. Cohen, 1967. Starch level in the trunk as a measure of compatibility between stock and scion in citrus. J. Hortic. Sci., 42: 331-334.
CrossRefDirect Link - Santamour, Jr. F.C., A.J. McArdle and R.A. Jaynes, 1986. Cambial isoperoxidase patterns in castanea. J. Environ. Hortic., 4: 14-16.
PubMed - Zapata, C., E. Deleens, S. Chailou and C. Magne, 2004. Partitation and mobilization of starch and N reserves in grapevine (Vitis vinifera L.). J. Plant Physiol., 161: 1031-1040.
PubMed