
Irving V. Occena
Institute of Chemistry, University of the Philippines Los Banos, College, Laguna, Philippines
Elmer-Rico E. Mojica
Institute of Chemistry, University of the Philippines Los Banos, College, Laguna, Philippines
Florinia E. Merca
Institute of Chemistry, University of the Philippines Los Banos, College, Laguna, Philippines
Asian Journal of Plant Sciences
Year: 2007 | Volume: 6 | Issue: 5 | Page No.: 757-764







ABSTRACT
The aim of this study is to isolate lectin from a seed which had no economical use. A lectin was isolated and purified from the seeds of Artocarpus camansi Blanco by extraction with phosphate buffered saline at pH 7.2. Further purification was done by precipitation with ammonium sulfate at 0-90% saturation. The dialyzed ammonium sulfate precipitate was subjected to gel filtration chromatography using Sephadex G-200 and fractions were tested for lectin activity. Native PAGE was employed to establish the homogeneity of the lectin positive fractions. Fractions with the same band were pooled and lyophilized. The purified lectin is non-blood type specific since it agglutinated all human blood types. It is blood group specific because it agglutinated blood from cow but not from swine. The purified lectin is a complete lectin since it agglutinated erythrocytes without trypsin treatment and addition of metal ions. However, trypsination of erythrocytes increases the agglutination activity. The isolated lectin did not show any specificity towards the ten sugars used in this study. The molecular weight of the purified lectin was estimated by using Sephadex G-200 and was found to be around 2,000 kDa since it eluted in the void volume. The lectin was found to be stable at 20-40°C.
PDF Abstract XML References Citation
How to cite this article
Irving V. Occena, Elmer-Rico E. Mojica and Florinia E. Merca, 2007. Isolation and Partial Characterization of a Lectin from the Seeds of Artocarpus camansi Blanco. Asian Journal of Plant Sciences, 6: 757-764.
DOI: 10.3923/ajps.2007.757.764
URL: https://scialert.net/abstract/?doi=ajps.2007.757.764
DOI: 10.3923/ajps.2007.757.764
URL: https://scialert.net/abstract/?doi=ajps.2007.757.764
INTRODUCTION
Lectins are divalent or multivalent carbohydrate-binding proteins that can agglutinate cells or other materials that contain more than one saccharide or sufficient complementary sites (Barondes, 1981). They may also be glycoproteins that can agglutinate human and/or animal erythrocytes and can also precipitate polysaccharides and complex arbohydrates (Goldstein et al., 1980). They are originally called phytohemagglutinins since they were first found in plants and later termed as agglutinins or hemagglutinins. Studies have proven that lectins can be found in various sources-plants, animals and even lower form of organisms. The most abundant lectins can be found in the seeds of leguminous plants but they can also be obtained from other plant parts (leaves, roots and stems).
Lectins are used as probes for identifying and mapping sugars on cell surfaces and as specific reagent in affinity chromatography for the isolation of saccharide-containing biomolecules because of its agglutinating property and their ability to bind glycoconjugates (Sharon, 1977). These properties are also the basis in using lectins for blood typing, cell separation, bacterial typing and bone marrow transplantation (Lis and Sharon, 1986). It can also be a tool in cancer research due to its ability to differentiate malignant (or transformed) cells from the normal ones (Smetana et al., 2006). Lately, lectins have been used as biotechnological tools in different studies (Kabir, 1998; Amadeo et al., 2003; Texeira et al., 2004; Ganiko et al., 2005).
Another possible function of lectins is their participation in binding nitrogen-fixing bacteria to legume roots. Studies have shown that lectins play an important role in the mechanism of binding Rhizobium to root hairs of legume plants (Barondes, 1981). Because of the various applications and uses of lectins, it is of utmost importance to isolate and purify lectins from abundant and less expensive sources.
The camansi (Artocarpus camansi Blanco) is the seeded form that belongs to the Moraceae family while its seedless counterpart is called rimas in Tagalog (Artocarpus altilis). The tree usually reaches up to 25 m high, with very large pinnate leaves (Pancho, 1979). In the Philippines, its fruit is used as food, cooked with coconut milk. The tree is used as windbreak and is also multi cropped with coffee. They usually thrive in tropical countries and are abundant in the Philippines and in other islands of the Pacific (Verheij and Coronel, 1980). The general objective of this study is to isolate and partially characterize the lectin in Artocarpus camansi Blanco seeds.
MATERIALS AND METHODS
Collection of samples: Seeds extracts from different plant sources were assayed for the presence of lectins. The plant species used were: Citrus grandis (pomelo), Citrus sinensis (orange), Mangifera indica (mango), Artocarpus camansi (camansi), Carica papaya (papaya), Citrofortunella microcarpa (calamansi), Durio zibethinus (durian), Malus sylvestris (apple) and Citrullus lanatus (watermelon). The seeds were collected from Tagum City, Davao Philippines. The collection of samples was done in 2005 and the other part of the experiments was conducted at the Institute of Chemistry, UPLB, Laguna, Philippines after the collection of samples.
Preparation of crude extracts: The dried seed samples were dehulled manually and ground using mortar and pestle, then defatted using n-hexane at ratio of 1:5 w/v for 15 min. Defatting was done twice until the hexane was clear of fat. 0.02 M PBS (pH 7.2) was added to the air-dried sample at a ratio of 5:1 (v/w) then subjected to continuous stirring using a magnetic stirrer for 16 h. After stirring, the sample was filtered through mira cloth. The liquid portion was clarified by centrifugation at 9000xg for 45 min at 4°C. After centrifugation, the residue was discarded and the clarified crude extract was stored at 0°C for further use. Before the purification process, the crude extract was subjected to hemagglutination assay and protein determination.
Ammonium sulfate precipitation: The crude extract was subjected to 90% ammonium sulfate precipitation using Cooper’s nomogram (1977). For every 100 mL crude extract, 61.1 g of ammonium sulfate was added in three lots (with stirring) for 30 min. After standing for 1 h, the mixture was subjected to 9000xg centrifugation and the precipitate collected were dissolved in minimum amount of buffer and stored at 0°C for further use. Addition of ammonium sulfate, standing and centrifugation procedures were performed at 4°C to prevent protein denaturation.
Desalting: The ammonium sulfate in the previous procedure was removed using a Sigma membrane dialysis tubing with molecular weight cutoff of 12 kDa. The tubing was pretreated by subjecting it to running water for 3-4 h to remove the glycerin which acts as humectant. The tube was sealed at one end with a cotton string then the ammonium sulfate fraction was introduced through the other opening with the use of a pipette to ensure no formation of bubbles. After sealing the other end, the tube was placed in a beaker filled with distilled water and stirred for 6 h at 4°C with three changes of water. The desalted sample was collected by punching a hole to one end, allowing the liquid to flow in a container. The dialyzed sample was stored at 0°C for further use.
Gel permeation chromatography: Purification of the 0-90% ammonium sulfate dialyzed fraction was done by using Sephadex G-200 column with dimension 27x0.4 cm and bed volume of 13.5 cm3. A 0.02 M PBS (pH 7.2) was used to equilibrate and elute the column. After reading the absorbance at 280 nm, the eluted fractions of 3.0mL were assayed for lectin activity by hemagglutination assay. All fractions that exhibited agglutination were pooled and concentrated using polyethylene glycol (PEG) and freeze-dried for later use.
Protein determination: The dye-binding method of Bradford (1976) was used in determining the protein content of the crude extract, ammonium sulfate fractions and the concentrated Sephadex G-200 fractions that showed lectin activity. The standard used was bovine serum albumin (BSA). About 1.5 mL of coomassie blue reagent was added to 0.5 mL of the sample then mixed thoroughly. The absorbance of the mixture at 595 nm using DU Beckman 650 UV-VIS spectrophotometer was recorded after standing for 5 min.
Preparation of red blood cells: A 200 μL blood (slightly outdated) of different types (A, B, AB, O) was mixed with 10 mL Phosphate Buffer Saline (PBS) with pH 7.2 in a centrifuge tube. The mixture was then subjected to a 5 min centrifugation at 1000xg. The centrifugate was removed and discarded using a Pasteur pipette. The compacted red blood cells were diluted with PBS until the volume reaches the 10 mL mark. Dilution and centrifugation was repeated until the supernatant is clear. The erythrocytes were diluted to 10 mL mark using PBS to make a 2% blood suspension.
For trypsination process, 1% trypsin was added to 2% blood suspension at a ratio of 1:1 (v/v) and was incubated at 37°C. After 1 h, the trypsinized cells were washed with PBS three times. Diluting 9.7 mL of PBS to 0.15 mL of packed cells made a 1.5% blood suspension.
Hemagglutination assay: Fifty microliters of samples (crude extract, ammonium sulfate fractions and gel filtration fractions) were serially two-fold diluted using PBS in plastic multi-well microtiter plates. Then 50 μL of 2% (v/v) erythrocyte suspension were added in each well. The bloods used were of human blood as well as trypsinized and untrypsinized animal blood (cow and swine).
After an incubation period of one hour, the plates were examined visually. A uniform layer of the erythrocytes over the surface of the well indicates positive agglutination. Negative result was indicated by the formation of a distinct button at the bottom of the well. The reciprocal of the maximal dilution of extract that exhibited positive reaction is the agglutination titer. Specific activity of the lectin was calculated by dividing the titer by the protein content, which is expressed in mg per mL extract.
Polyacrylamide gel electrophoresis: The crude extract, ammonium sulfate fraction and the pooled Sephadex G-200 fractions were subjected to native PAGE (polyacrylamide gel electrophoresis) to determine the purity of the samples. Native PAGE was carried out in 5% polyacrylamide stacking gel and 10% polyacrylamide separating gel in Tris-HCl buffer pH 8.8 following the procedure by Laemmli (1970). The tracking dye used in the electrophoresis was Sucrose Bromphenol Blue (SBB) and the tank buffer consisted of 0.025 M tris and 0.192 M glycine at pH 8.3. Electrophoresis was carried out at 70 Volts and 15 mA initial current. When the tracking dye reaches the separating gel, the current was increased to 20 mA. Silver staining was used for detection of the bands.
Molecular weight estimation by gel filtration chromatography: The molecular weight of the isolated lectin was determined using gel filtration chromatography. The void volume of a G-200 column was determined by running blue dextran and determining the absorbance of the fractions (3 mL per tube) at 595 nm. A mixture of standards was loaded to the same gel and the absorbance of each tube was obtained at the same wavelength. The relative elution volumes of the standards were plotted against the logarithm of their molecular weights to construct a calibration curve. From this curve, the molecular weight of the purified lectin was extrapolated. The following standards were used in constructing the calibration curve: cytochrome C, 12.4 kDa; carbonic anhydrase, 29 kDa; bovine serum albumin, 66 kDa, alcohol dehydrogenase, 150 kDa and β-amylase, 200 kDa.
Inhibition of agglutination: The following sugars were used to determine its effect on the agglutination reaction: glucosamine, glucose, maltose, lactose, galactose, arabinose, mannose, fructose, xylose, rhamnose, raffinose and fucose.
The concentrations of the standard sugars used were 1000 and 500 mM. A 50 μL of the isolated lectin was placed in multiwell microtiter plates that contained 50 μL of the sugars mentioned above. Inhibition of agglutination was observed after adding 50 μL of 2% (v/v) erythrocyte suspension and the formation of a distinct button of red blood cells at the bottom of the well.
Effect of temperature on lectin activity: The samples were first incubated in a water bath for 30 min at 20, 30, 40, 50, 60, 70, 80 and 90°C. Hemagglutination assay was then performed for each sample.
RESULTS AND DISCUSSION
Isolation and purification of lectin: In the initial survey for the presence of lectins from the seeds of different plant species, only blood type O was used for hemagglutination assay and the result of the survey was presented in Table 1. This survey only employs 4 h of extraction with PBS and no defatting was done. Durian (Durio zibethinus) neither show any agglutination nor formed a distinct button of erythrocytes at the bottom of the well (which would indicate the absence of agglutination). Durian seed is rich in fat and that may have been the interfering factor in the agglutination since fat has been found to inhibit agglutination (De Mesa et al., 2004). Among the 9 plant species tested, the seed from Artocarpus camansi plant (Fig. 1), a member of the Moraceae plants, has the highest titer value (2.048) indicating high lectin content. This is the reason why camansi seeds were chosen as the subject of this study.
The detection of presence of lectin in the sample used added another Artocarpus species from which lectin can be isolated. The most popular lectin, jacalin have been isolated from seeds of jackfruit (Artocarpus heterophyllus) and is one of the most studied lectin (Kabir, 1995; Gaikwad, 2006). Another lectin known as heterophyllin was also isolated from A. heterophyllus seeds (Mourao et al., 1999). Other lectins isolated from the seeds of Artocarpus species and Moraceae family included seeds from A. altilis Humter (1986), A. lakoocha (Chowdhury et al., 1987), A. hirsuta (Antony et al., 1989), A. integer (Lim et al., 1997) A. incisa (Moreira et al., 1998), A. lignanensis (Zhang et al., 1999), A. integrifolia (Silva-Lucca et al., 1999) and lately two species from Vietnam namely A. champeden and A. masticata (Tran et al., 2006).
| Table 1: | Survey for the presence of lectins from the seeds of different plant sources |
 | |
| 1Titer value is the reciprocal of the maximal dilution of lectin which can cause agglutination | |
 | |
| Fig. 1: | The hulled and dehulled seeds of Artocarpus camansi Blanco seeds |
The dehulled, ground seeds were first extracted with n-hexane to remove fats or lipids that may interfere with agglutination. In the preliminary studies, there were no observed differences in lectin activity in samples extracted with n-hexane compared to those not treated with the solvent. After extraction with n-hexane, the solvent remains clear. This indicates that the sample contains very little amount of lipid from the 60.5 g sample and 370 mL PBS, 335 mL of crude extract was obtained.
Proteins were separated from other substances co-extracted by PBS in the crude extract by precipitating it with ammonium sulfate. This process of protein precipitation is called salting-out where a salt solution decreases the solubility and eventually, precipitates the proteins. Salt solutions such as ammonium sulfate and magnesium sulfate absorbs the water of hydration surrounding and protecting the protein in the extract, causing the latter to coagulate. Ammonium sulfate was chosen as precipitating agent in this study due to its high solubility in water and produces high ionic strength. Increase in ionic strength decreases the protein solubility (Scopes, 1994). Zero to ninety percent (0-90%) ammonium sulfate saturation was employed to ensure that most proteins will be precipitated.
Ammonium sulfate was removed from the protein since it may cause false positive agglutination. Desalting was done by extensive dialysis with distilled water. Dialysis was performed in the cold room to prevent possible denaturation of the protein. The membrane used has a molecular cut-off of 12 kDa. This allows the proteins with molecular weight greater than 12 kDa to remain inside the dialysis bag.
Gel filtration chromatography using Sephadex G-200 can resolve proteins with molecular weights up to 600 kDa. Figure 2 shows the elution profile of the dialyzed 0-90% ammonium sulfate fraction at a flow rate of 0.1 mL per minute. Two peaks were visible after reading the absorbance of the 39 fractions collected (3 mL per tube) but only fractions 7-10 showed agglutination, which falls on the first peak. This may be an indication that the lectin is large and has high molecular weight since it was eluted first. Fractions collected corresponding to the second peak did not show any agglutinating activity. The fractions that showed positive test for lectin were pooled and lyophilized. The lyophilized lectin was subjected to native PAGE to determine its homogeneity. Fractions 7-10 showed a faint single band near the top of the separating gel that suggests that the purified sample contains only one lectin (Figure not shown).
Characterization of the purified lectin: Hemagglutination assay. The isolated lectin from A. camansi is considered to be non-blood type specific since it agglutinates all human blood types. This lectin could be similar to A. lignanensis lectin (Zhang et al., 1999) which is also non-blood type specific. However, it is a blood group specific lectin since it agglutinated blood from cow but not from swine (Table 2). Agglutination using animal blood was done only to these two groups due to unavailability of other animal blood. Since agglutination was not observed in swine blood, it suggests that the lectin is blood group specific.
The human blood types have different sugar moieties on the surface of the cell. Type A has N-acetyl-D-galactosamine, D-galactose for type B and L-fucose in type O. Blood type AB contains the sugar determinants for both A and B. Agglutination occurs when the lectin interacts with these sugar moieties. Non-blood type specificity of the lectin may be due to the presence of multiple binding sites where it can recognize all the determinants for each blood type (Aragones and Merca, 1998).
| Table 2: | Hemagglutination assay of purified A. camansi using animal blood |
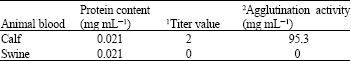 | |
| 1Titer value is the reciprocal of the maximal dilution of lectin which can cause agglutination; 2 Least concentration of lectin required to agglutinate 2% erythrocyte suspension | |
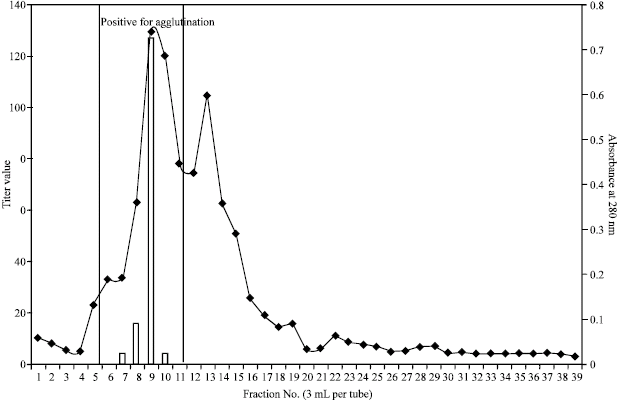 | |
| Fig. 2: | Elution profile and titer value of the 0-90% ammonium sulfate fractions on Sephadex G-200 |
| Table 3: | Effect of trypsin on agglutination using blood type B |
 | |
| 1Titer value is the reciprocal of the maximal dilution of lectin which can cause agglutination; 2Least concentration of lectin required to agglutinate 2% erythrocyte suspension | |
Trypsination of blood type B (Table 3) increased the agglutination of cells compared to untrypsinized blood. Trypsin probably redistributes the receptor sites in the erythrocyte’s surface, which induces clustering that is more favorable for agglutination (Goldstein and Hayes, 1978). The purified lectin is a complete lectin since it can agglutinate erythrocytes in saline solution without the aid of metal ions, trypsin or proteolytic enzymes.
The purified lectin was assayed for inhibition of agglutination using of 10 sugars. Even at 1000 mM sugar concentration, no inhibition was observed for all the sugars used. This may indicate that the lectin contains numerous binding sites that are compatible with a wide range of monosaccharide receptors. Inhibition would occur if the sugar-lectin interaction is stronger than the erythrocyte-lectin attraction and would consequently form a distinct button of the erythrocyte at the bottom of the well.
Non-inhibition of agglutination by the sugar molecules to the erythrocyte-lectin interaction may be due to the following reasons: (a) there is tolerance for substitution at certain positions of the sugar moiety which determines their specificity; (b) lectin binding sites are non-specific for sugar anomers; (c) size and shape of sugar residues does not affect the erythrocyte-lectin binding; and (d) the carbohydrate containing the sugar or its derivatives cross react with the lectin (Lacsamana and Merca, 1994). However, inhibition was tested only using 10 sugars so this may not be an enough reason to say that the A. camansi lectin is not sugar-specific.
Literature reported inhibition by monosaccharides on lectins like galactose and mannose on jacalin (Peumans et al., 2000) and mannose on arthocarpin (Barre et al., 2004) and galactose on A. lignanensis lectin (Zhang et al., 1999). Complex sugar like N-acetyl-D-glucosamine inhibited arthocarpin (Mourao et al., 1999) and A. lignanensis lectin (Zhang et al., 1999) while A. hirsuta lectin is inhibited by methyl-α-D-galactopyranoside, p-nitrophenyl (pNP)- α-D-galactopyranoside and pNP-α-D-N-acetylgalactosamine (Gurjar et al., 1998).
Literature have also reported the grouping of lectin-binding properties based on determinant structure rather than monosaccharide-inhibition pattern.
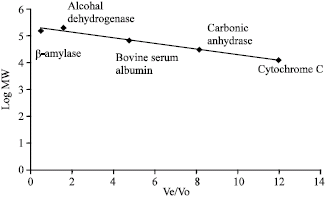 | |
| Fig. 3: | Calibration curve for the estimation of the molecular weight of the purified lectin from A. camansi seeds using gel filtration chromatography |
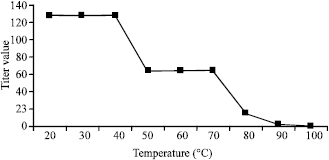 | |
| Fig. 4: | Effect of temperature on agglutination |
This was done to facilitate the selection of lectins as structural probes for glycans, as well as for the interpretation of the distribution and the properties of the carbohydrate chains on the cell surface. Lectins from Artocarpus species (jacalin and artocarpin) were classified as T-specific lectins (Wu and Shunji, 1991).
Molecular weight estimation: The molecular weight of the purified lectin was estimated with the use of Gel filtration chromatography (G-200). The void volume was first determined by running blue dextran in the column which was found out to be 15 mL. The calibration curve (Fig. 3) was constructed by running the different protein standards. It was found out that the molecular weight of the purified lectin is greater than or equal to 2,000 kDa since it eluted in the void volume.
The result is quite surprising since literature have shown that the molecular weight obtained from lectins isolated from other Artocarpus species are quite low. The jacalin from jackfruit have sub-units with molecular weight of 12 and 15 kDa (Kabir, 1995) while the heterophyllin has sub-units of 31.4, 18.7 and 16.3 kDa (Mourao et al., 1999). Molecular weight determined by gel filtration reported the lectin from A. hirsuta to have a molecular weight of 45 kDa (Antony et al., 1989), while A. incisa gave 48-49 kDa (Moreira et al., 1998). On the other hand, A. altilis have a molecular weight of 44.5 kDa using gel filtration with sub-units of 19 and 22 kDa. The highest molecular weight reported so far was that of A. lakoocha which is 70 kDa by gel filtration with four sub-units of 18 kDa (Chowdhury et al., 1987).
Effect of temperature: Blood type A was used to detect the presence of lectin in this characterization procedure since the purified lectin is observed to agglutinate all human blood types with the same titer value. The effect of temperature in agglutination is shown in Fig. 4. The lectin is stable from 20-40°C and has a consistent titer value of 128. At higher temperature the titer value decreased and at 100°C, it is devoid of lectin activity. This may be brought about by the denaturation of the lectin that removes its agglutinating capacity.
CONCLUSIONS
Lectin from the seeds of Artocarpus camansi Blanco was isolated and purified. The purified lectin was found to be non-blood type specific since it agglutinates all human blood types and blood-group specific to human and cow’s blood. The isolated lectin can be considered a complete lectin since it can readily agglutinate red blood cells without the further need of metal ions or proteolytic enzymes In addition, trypsination increased the agglutination of the red blood cells since there is an increase in titer value. The lectin was found to be stable at temperatures 20-40°C where it exhibited maximum agglutination.
REFERENCES
- Amadeo, G.I., R. Moreira, R. Lima, D. Teixeira, R. Kratje and M. Etcheverrigaray, 2003. Screening of lectins from South American plants used as affinity ligands to purify rhEPO. Braz. J. Chem. Eng., 20: 21-26.
CrossRefDirect Link - Aragones, R.C. and F.E. Merca, 1998. Two lectins from the seeds of lablab bean (Lablab purpureus Linn. cv. Highworth). Philipp. J. Sci., 127: 15-37.
Direct Link - Barre, A., W.J. Peumans, M. Rossignol, G. Borderies, R. Culerrier, E.J.M. Van Damme and P. Rouge, 2004. Artocarpin is a polyspecific jacalin-related lectin with a monosaccharide preference for mannose. Biochimie, 86: 685-691.
PubMedDirect Link - Bradford, M.M., 1976. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem., 72: 248-254.
CrossRefPubMedDirect Link - Gaikwad, S.M. and M.I. Khan, 2006. Binding of T-antigen disaccharides to Artocarpus hirsuta lectin and jacalin are energetically different. Photochem. Photobiol., 82: 1315-1318.
PubMedDirect Link - Ganiko, L., A.R. Martins, E. Freymuller, R.A. Mortara and M.C. Roque-Barreira, 2005. Lectin KM+-induced neutrophil haptotaxis involves binding to laminin. Biochim. Biophys. Acta, 1721: 152-163.
PubMedDirect Link - Goldstein, I.J. and C.E. Hayes, 1978. The lectins: Carbohydrate-binding proteins of plants and animals. Adv. Carbohydr. Chem. Biochem., 35: 127-340.
CrossRefDirect Link - Goldstein, I.J., R.C. Hughes, M. Monsigny, T. Osawa and N. Sharon, 1980. What should be called a lectin? Nature, 285: 66-66.
CrossRef - Kabir, S., 1995. The isolation and characterization of jacalin (Artocarpus heterophyllus (Jackfruit) lectin] based on its charge properties. Int. J. Biochem. Cell Biol., 27: 147-156.
Direct Link - Kabir, S., 1998. Jacalin: A jackfruit (Artocarpus heterophyllus) seed-derived lectin of versatile applications in immunobiological research. J. Immunol. Methods, 212: 193-211.
CrossRefPubMedDirect Link - Laemmli, U.K., 1970. Cleavage of structural proteins during the assembly of the head of bacteriophage T4. Nature, 227: 680-685.
CrossRefDirect Link - Lim, S.B., C.T. Chua and O.H. Hashim, 1997. Isolation of a mannose-binding and IgE- and IgM-reactive lectin from the seeds of Artocarpus integer. J. Immun. Methods, 209: 177-186.
PubMedDirect Link - Lis, H. and N. Sharon, 1986. Lectins as molecules and as tools. Annu. Rev. Biochem., 55: 35-67.
CrossRefDirect Link - Moreira, R.A., C.C. Castelo-Branco, A.C.O. Monteiro, R.O. Tavares and L.M. Beltramini, 1998. Isolation and partial characterization of a lectin from Artocarpus incisa L. seeds. Phytochemistry, 47: 1183-1188.
CrossRefDirect Link - Mourao, R.H.V., F.J. Xavier, E.W. Alves, E.G. Orellano and D. Fernandes de Melo et al., 1999. Isolation and partial characterization of heterophyllin, a new lectin from Artocarpus heterophyllus seeds. Acta Farmaceutica Bonaerense, 18: 41-47.
Direct Link - Peumans, W.J., B. Hause and E.J.M. Van Damme, 2000. The galactose-binding and mannose-binding jacalin-related lectins are located in different sub-cellular compartments. FEBS Lett., 477: 186-192.
Direct Link - Silva-Lucca, R.A., M. Tabak, O.R. Nascimento, M.C. Roque-Barreira and L.M. Beltramini, 1999. Structural and thermodynamic studies of KM+, a D-mannose binding lectin from Artocarpus integrifolia seeds. Biophys. Chem., 79: 81-93.
PubMedDirect Link - Smetana, K. Jr. B. Dvorankova, M. Chovanec, J. Boucek and J. Klima et al., 2006. Nuclear presence of adhesion-growth-regulatory galectins in normal/ malignant cells of squamous epithelial origin. Histochem. Cell Biol., 125: 171-182.
CrossRefDirect Link - Teixeira, S.A., M.S. Viapiano, L. Ganiko, M.C. Roque-Barreira and A.R. Martins, 2004. The novel lectin KM+ detects a specific subset of mannosyl-glycoconjugates in the rat cerebellum. Glycoconj. J., 20: 501-508.
PubMedDirect Link - Tran, T.P.L., V.H. Nguyen, T.Q. Nguyen and N.L. Do, 2006. Comparison of the serum IgA1 antibody capturing properties of lectins isolated from three jackfruit species of Vietnam: Artocarpus champeden (Mit to nu), A. heterophyllus Lamk (mit dai) and A. masticata (mit chay). Tap. Chi. Duoc. Hoc., 46: 16-18.