
Agbagwa Ikechukwu Ozoemenam
Department of Plant Science and Biotechnology, Faculty of Science, University of Port Harcourt, Nigeria
Okoli Bosa Ebenezar
Department of Plant Science and Biotechnology, Faculty of Science, University of Port Harcourt, Nigeria
Ndukwu Benjamin Chinyem
Department of Plant Science and Biotechnology, Faculty of Science, University of Port Harcourt, Nigeria
Asian Journal of Plant Sciences
Year: 2007 | Volume: 6 | Issue: 5 | Page No.: 732-740







ABSTRACT
Comparative anatomical studies of three Abrus Adanson species (A. precatorius, A. pulchellus and A. canescens) and a new collection identified as Abrus sp. were undertaken by simple microtomy. Parts of each species (seed, stem, root, leaf, pulvinus and petiole) were sectioned after fixation and wax embedding with a LEITZ 1512 rotary microtome at 20 to 24 μm thickness and observed with a LEITZ DIAPLAN photomicroscope. The seed coat in the species possesses five distinct layers, which varied in thickness. Similarities were observed in the arrangement, differentiation and distribution of cells and tissues in the investigated organs with characteristic pubescence in all organs of A. canescens. However, the number of layers of these cells and tissues (collenchyma, pericyclic parenchyma, sclerenchyma, pith parenchyma and the vascular cells) occasionally differed and could be harnessed for taxonomic purposes. The study aims at providing detailed comparative anatomical information for the genus, which is hitherto unavailable among the West African species. These will complement the morphological descriptions, which always overlap.
PDF Abstract XML References Citation
How to cite this article
Agbagwa Ikechukwu Ozoemenam, Okoli Bosa Ebenezar and Ndukwu Benjamin Chinyem, 2007. Comparative Anatomy of Abrus Adanson Species in Parts of Tropical West Africa. Asian Journal of Plant Sciences, 6: 732-740.
DOI: 10.3923/ajps.2007.732.740
URL: https://scialert.net/abstract/?doi=ajps.2007.732.740
DOI: 10.3923/ajps.2007.732.740
URL: https://scialert.net/abstract/?doi=ajps.2007.732.740
INTRODUCTION
Abrus Adanson is a member of the tribe Vicieae in the sub-family Papilionoideae of the Leguminosae. In West Africa Hutchinson and Dalziel (1958) and Burkill (1995) briefly described three species, which occur in Nigeria. These are A. canescens Welw. ex Bak., A. pulchellus Wall. ex Thw. and the widespread A. precatorius L. The species are climbing, twining or scrambling slender sub-woody liana with pinnate leaves, rachis ending in a bristle, stamens connate in a sheath with racemose flowers. Medicinal, pharmacological and toxicological properties and uses are ascribed to the species (Bouquet and Debray, 1974; Burkill, 1995). The vegetative parts (leaves and stems) of the species are very variable and plastic (Agbagwa and Okoli, 2005a, 2006), showing marked overlap in their characteristics, thereby making their usage in species identification difficult. This is exacerbated by the fact that floral characters used by Hutchinson and Dalziel (1958) to describe the two common species, A. precatorius and A. pulchelus, did not match with fresh collections made in Nigeria between 1998 and 2002. Hutchinson and Dalziel (1958) descriptions were probably based on herbarium specimens, which might not have been properly documented by the initial collectors. Also we were unable to make fresh collections of A. canescens in the course of this study after several field trips; thus only herbarium samples from Forestry Herbarium Ibadan (FHI) and University of Ibadan Herbarium (UIH) were used. The re-emergence of medicinal plants as healthcare alternatives (Hoareau and DaSilva, 1999) calls for greater interest into poorly utilized plant genetic resources as these. In the face of increasing threat to these resources (Abrus species exist in jungles, galleried forests and sacred groves, which are fast disappearing due to deforestation), any effort at better understanding and proper documentation of the individual species is worthwhile.
There have been recent taxonomic treatments of the genus Abrus dealing with micromorphology (Teixeira and Diniz, 2003; Agbagwa and Okoli, 2005a, 2006). Apart from Cutler (1978) who gave an anatomical overview of the stem of A. precatorius, no other known report on the anatomy of Abrus exists to the knowledge of the authors. The internal parts and structure of plants was suggested by Stace (1980) to be useful in the taxonomic delimitation of plants since they are less affected by environmental changes and therefore, highly conservative in taxonomic variations. The deposition of sclerenchyma, arrangement and formation of vascular bundles, differentiation of epidermal long and short hairs and other anatomical characters have been reported and utilized at different systematic levels for taxa elucidation or applied to resolve taxonomic conflicts (Metcalfe and Chalk, 1979; Stace, 1980; Kiew and Ibrahim, 1981; Fahn, 1990; Ndukwu and Okoli, 1992; Agbagwa and Ndukwu, 2004). Comparative and systematic studies on the anatomy of the leaf (including petiole and pulvinus), seed coat, stem and root of species of Abrus occurring in tropical West Africa is being presented for the first time. This study is aimed at providing a description of the anatomical features of the individual species on one hand and making anatomical comparison between the species for the purposes of taxonomic delimitation. The anatomical identity of a new collection, which is undergoing confirmation and simply referred to in this study as Abrus sp. is also presented. Systematic information on this medicinal genus is scanty with species facing extinction (Abrus species exist in jungles, galleried forests and sacred groves, which are fast disappearing due to deforestation), it is hoped that the overall results of the study will help in filling the gap in their taxonomy and stimulate interest in further studies leading to their conservation.
MATERIALS AND METHODS
The study was carried out in the Taxonomy and Biosystematics Research Lab, Department of Plant Science and Biotechnology, University of Port Harcourt.
Fresh materials collected during field trips and preserved materials from Forestry Herbarium Ibadan (FHI) and University of Ibadan Herbarium (UIH) were used (Table 1).
Fresh seed, stem, leaves, petiole and pulvinus of A. precatorius, A. pulchellus and Abrus sp. were fixed in Formal-Acetic-Alcohol (FAA) for 48 h, washed in several changes of distilled water, dehydrated through alcohol series (30, 50, 70, 95 and 100%), 2 h in each solution and embedded in wax. All samples of A. canescens used were herbarium materials. These were first soaked in water for upwards of 24 h to revive the samples before fixing in FAA for 48 h. Sections in each case were cut on a Leitz 1512 rotary microtome at thickness between 20 and 24 μm. The sections were dewaxed with pure xylene and rehydrated in alcohol series following Cutler (1978) with modifications. Staining was achieved by dipping the slides in 1% alcian blue for about 5 min, washed with distilled water and counter stained with 1% safranin for 2 min. The stained sections were dehydrated through alcohol series and mounted permanently in DPX (Dee Pex). Photomicrographs of the anatomical sections were taken with a Leitz Diaplan photomicroscope fitted with Leica WILD MPS 52 camera at X10 microscopic objective lens.
| Table 1: | Sources of plant materials used for the study |
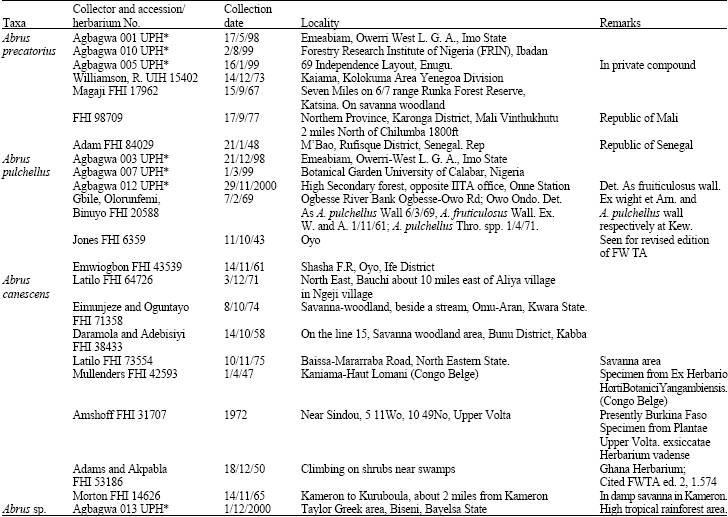 | |
| UPH*: Fresh specimens deposited at the University of Port Harcourt Herbarium | |
Microscope measurements of the sizes of cells and tissues in stem, leaf and seed were made at X10 microscope objective lens following Radford et al. (1974). Samples of the same age were used for the comparative studies in A. precatorius and A. pulchellus.
RESULTS
Observations and measurements made on the species are presented in Table 2-4 while Fig. 1-6 are photomicrographs of the various anatomical sections.
Stem: The same tissue types were observed in the species with slight differences in layers and arrangement (Fig. 1-5 and Table 2-4). The single-layered stem epidermis in A. precatorius is followed by angular collenchyma, sclerenchyma (brachysclereids), parenchyma (pericycle) and the vascular bundle. In A. pulchellus, the stem (Fig. 1b) epidermis is replaced by 3-5 layers of columnar shaped periderm or cork cells. The single-layered stem epidermis observed in A. canescens is piliferous and preceded by 2-3 layers of angular collenchyma. In the new collection Abrus sp. (Fig. 1c), the stem possesses a thick layer of outer cuticle followed by a single layer of flat epidermal cells. Stele in all species is siphonostele.
| Table 2: | Summary of Stem Anatomical features in Abrus species |
 | |
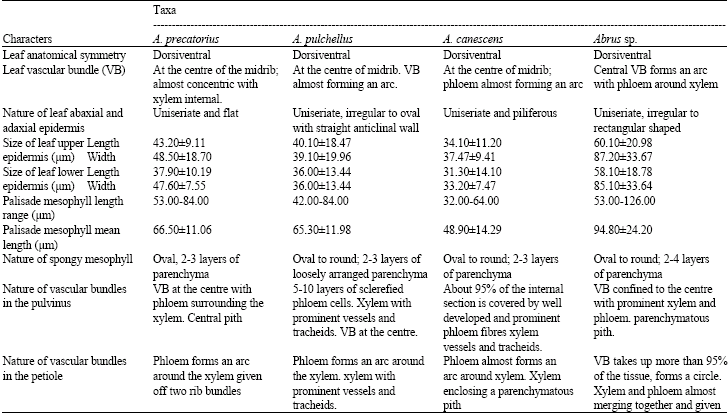
| Table 3: | Summary of anatomical features of the leaf, pulvinus and petiole of Abrus species |
 | |
| Table 4: | Summary of mean thickness of the seed coat (mm) of Abrus species studied |
 | |
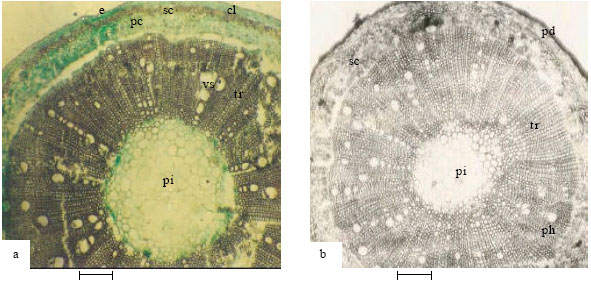 | |
| Fig. 1: | Showing stem anatomy of Abrus species (1a-A. precatorius; 1b-A. pulchellus; 1c-Abrus sp.). e-epidermis, cl-collenchyma, sc-sclerenchyma, pc-pericyclic parenchyma, pi-pith, tr-tracheids, vs-vessels, pd-periderm, ph-phloem (Bar: 100 μm) |

Root: Differences were observed in the number of layers of tissues. In A. precatorius, the root anatomy (Fig. 2a) begins with a thick layer of cork followed by 3 cell layers of irregularly shaped and thick walled periderm cells, 3-4 layers of cortical cells and 1-2 layers, thick walled endodermal cells with rays. Several layers of sclerified phloem fibres are buried in the tissues with some obliterating the endodermis. Xylem occurs towards the centre. The pith is characterized by sclerified parenchyma and some metaxylem cells. In A. pulchellus, the root anatomy (Fig. 2b) consists of an outer layer of cork cells followed by 5 cell layers of compactly arranged, isodiametrical and rectangular periderm cells, 3 layers of cortical cells, 1-2 layers of thick walled endodermal cells with rays. Phloem fibres occur in clusters around the phloem area; 2-22 cell clusters of phloem fibres occasionally obliterate the endodermis.
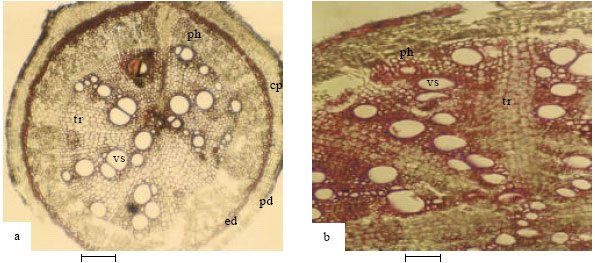 | |
| Fig. 2: | Showing root anatomy of Abrus species (2a-A. precatorius; 2b-A. pulchellus). cp-cortical parenchyma, ed-endodermis (Bar: 100 µm) |
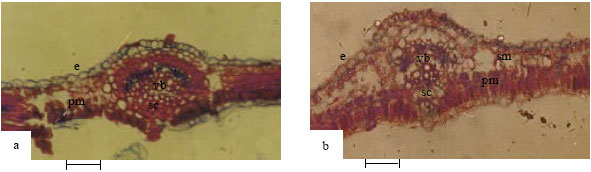 | |
| Fig. 3: | Showing cross-section of leaf of Abrus species (3a-A. canescens; 3b-A. pulchellus). e-epidermis, pm-palisade mesophyll, sm-spongy mesophyll, sc-sclerenchyma, vb-vascular bundle (Bar: 65 µm) |
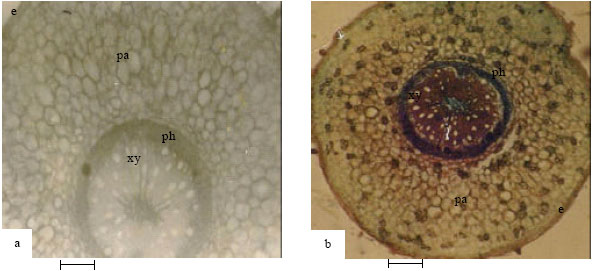 | |
| Fig. 4: | Showing cross-section of pulvinus of Abrus species (4a-A. pulchellus, 4b-A. precatorius). e-epidermis, pa-parenchyma, ph-phloem, xylem (Bar: 100 µm) |
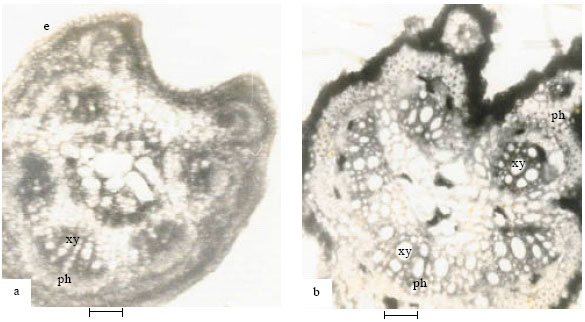 | |
| Fig. 5: | Showing cross-section of petiole of Abrus species (5a-A. precatorius, 5b-A. canescens). xy-xylem, ph-phloem (Bar: 100 μm) |
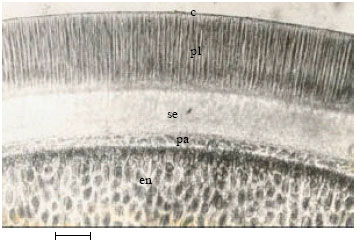 | |
| Fig. 6: | Showing cross-section of seed coat of Abrus species (A. precatorius). c-cuticle, pl-palisade (macrosclereid) layer, se-supepidermis (lagenosclereid), pa-seed parenchyma, en-endosperm (Bar: 120 μm) |
In the new collection Abrus sp., 5 layers of periderm cells, 4 to 6 layers of cortical cells and 2 to 3 layers of large and thick walled endodermal cells with rays were observed.
Leaf: Observations on the leaf anatomy (Fig. 3a and b) show that the three species and the new collection are dorsiventral, with uniseriate and irregularly shaped epidermal cells. The epidermis is uniseriate in A. canescens. Two layers of palisade mesophyll cells were observed in all species with those in A. precatorius open in arrangement. Apart from in the new collection Abrus sp., where 2-4 layers of spongy mesophyll cells were observed, others had 2-3 layers. Collenchyma and sclerenchyma layers around the vascular bundles differed: 2-4 layers in A. precatorius, 2-5 layers in A. pulchellus, 2-4 layers in A. canescens and 2-9 layers in the new collection Abrus sp.
Pulvinus and petiole: The pulvinus in A. precatorius (Fig. 4a and Table 3) consists of a single layer of occasionally raised and cutinized epidermis followed by 4-7 layers of parenchyma and a central vascular bundle. Pith is present. The vascular bundle of the pulvinus continues into the petiole (Fig. 5a and Table 3). The petiole also has a layer of round to ovoid epidermal cells and 4-6 layers of collenchyma. In A. pulchellus, the pulvinus (Fig. 4b) has a single layer of isodiametric, flat and cutinized piliferous epidermis preceded by 9-10 layers of thin-walled parenchyma cells. 1-3 layers of collenchyma cells surround the vascular bundle. Pith is present. The petiole possesses a layer of irregularly shaped, round to ovoid epidermal cells, 3-4 layers of parenchyma and collenchyma cells. Xylem and phloem are separated by interfascicular cambium.
In A. canescens, the vascular system of the pulvinus pushes a few layers of parenchyma and collenchyma cells close to the epidermis. The single piliferous cutinized epidermis of the petiole (Fig. 5b) is followed by 2-3 layers of parenchyma. In the new collection Abrus sp., the petiole has a single layer of cutinized epidermis followed by 2-4 layers of collenchyma.
Seed coat: The seed-coat (Fig. 6) in all the species have 5 tissue layers; cuticle, followed by a palisade layer of long macrosclereids, a sub-epidermal layer of columnar to flask-shaped lagenosclereids, chlorenchymatous parenchyma and finally an area of aleurone endosperm.
DISCUSSION
The evidence from anatomical studies shows that variations exist among the species of Abrus studied. The taxonomic value of such variations in anatomical features between species has been reported (Stace, 1965, 1980; Tateoka, 1969; Carlquist, 1961; Esau, 1977; Metcalfe and Chalk, 1979; Ndukwu and Okoli, 1992; Agbagwa, 2001; Agbagwa and Ndukwu, 2004).
The stem anatomy of A. precatorius, A. pulchellus and A. canescens were observed to be similar (Table 2, Fig. 1a and 1b) to the outline description of Cutler (1978) for A. precatorius. A major difference is the replacement of the epidermis in A. pulchellus by 3-5 layers of columnar shaped peridermal or cork cells. Other variations were observed to be due to the number of layers of certain tissues like collenchyma, pericyclic parenchyma, sclerenchyma and pith cells. The number of tracheids and the large solitary vessels occurring in radial chains, which are similar to those reported by Cutler (1978), varied between species. A distinct anatomical feature of A. canescens stem is the possession of epidermal hairs and degeneration of the pith cell pushing outwards the parenchymatous pith cells and leaving a prominent central hollow. Whether this is an adaptive anatomical modification or a phylogenetic advancement is subject to confirmation. Though the anatomy of the new Abrus sp. is generally similar to the other species, it characteristically has well cutinized epidermis and a 3-layered parenchymatous region between the collenchyma and scherenchyma instead of between the sclerenchyma and the vascular regions as was observed in the other species. This re-arrangement demonstrates the relevance of these anatomical features in taxonomic delimitation of the species.
Comparative anatomy of the leaf, petiole and pulvinus of the species revealed similarity and overlap in certain features. Though similarity of characters where they occur, connote relatedness of species, observed differences in vascular bundle arrangement, number of layers of sclerotic cells surrounding the vascular bundles, number of layers of parenchyma and collenchyma cells, length of the palisade cells and thickness of upper and lower leaf epidermis, offer useful parameters for delimiting the various taxa in this genus. Such anatomical differences have been previously utilized in taxa elucidation (Agbagwa and Ndukwu, 2004). For instance, the piliferous epidermis of the stem, leaf, petiole and pulvinus of A. canescens, a feature observed only in this species, distinguishes it from the other species. It was also observed that the upper and lower leaf epidermal cells of the species varied in thickness. The new collection Abrus sp. (60.10 ± 20.98x87.20 ± 33.67 μm for upper epidermis and 58.10 ± 18.78x85.10 ± 33.64 μm for lower epidermis) had the highest values for leaf epidermal thickness. In all the species the upper leaf epidermal layers were thicker than the lower epidermises; a feature associated with the greater exposure of the upper epidermis to sunlight and the need to guard against water loss (Esau, 1977; Chen and Wen, 2005; Agbagwa and Okoli, 2006). While the longest palisade cells were observed in Abrus sp., the closeness of length of palisade mesophyll cells in A. precatorius and A. pulchellus is noteworthy. The leaf vascular bundles of A. canescens and Abrus sp. are quite similar with the phloem forming an arc around the xylem. Thus there is an obvious variation in affinities of taxonomic features within species.
The arrangement of tissues in the pulvinus of the four species is the same and uniquely conforms to the description given by Esau (1977) on Mimosa pudica L., another legume. It is possible that this arrangement of cells is a characteristic of pinnate members of the leguminosae that respond to touch stimuli. The petiole anatomy of the species, which is characterized by the phloem forming an arc around the xylem and giving off 2 rib bundles at the periphery, differed in the new collection Abrus sp. The vascular systems of Abrus sp. almost form a central circle with seven vascular bundles. The cambium in each group separates the xylem innermost and the phloem outermost. This difference in the arrangement of the petiolar vascular bundles can be exploited at the species level for the separation of the new collection Abrus sp.
The seed coat anatomy of the four species (Fig. 6 and Table 4) only differed in the thickness of the five identified layers. The value of such differences in thickness in taxa elucidation has been highlighted by Esau (1977) using Crotolaria intermidia a leguminous plant. The fact that the thickness of the cuticle, palisade and subepidermal layers were all highest in A. precatorius (Table 3) may be connected with the very hard, tough and impenetrable seeds of the species (Agbagwa, 2001). The hard seed coat may have developed from the deposition of several cuticular materials and presence of special types of sclerotic cells. These features confer hardness on this species and thus separate its seeds from the other Abrus species.
The observed anatomical similarities among the Abrus species studied indicate phyllogenetic relatedness of the taxa. Although, Bailey (1951, 1953 and 1957) had suggested that similarities in structural specialization do not necessarily imply close relationship but may be the result of parallel and convergent evolution. However, previous studies involving other lines of taxonomic evidence (Agbagwa, 2001; Agbagwa and Okoli, 2005a, b, 2006) clearly indicate that the species of Abrus studied belong to different taxa. Therefore, the anatomical differences observed in each species must have evolved with that particular species to confer heritable variation that could be exploited for taxonomic purposes. The current study is an attempt to provide anatomical data on the three Abrus species earlier described by Hutchinson and Dalziel (1958) and the new collection from southern Nigeria. These will complement the morphological descriptions, which always overlap.
ACKNOWLEDGMENT
We are grateful to the Management and Staff of Forestry Herbarium Ibadan and the Curator at the University of Ibadan Herbarium for their assistance during the study.
REFERENCES
- Agbagwa, I.O. and B.C. Ndukwu, 2004. The value of morpho-anatomical features in the systematics of Cucurbita L. (Cucurbitaceae) species in Nigeria. Afr. J. Biotechnol., 3: 541-546.
Direct Link - Agbagwa, I.O. and B.E. Okoli, 2005. Fruit epidermal micromorphology in the systematics of Abrus Adanson (Papilionaceae) in parts of West Africa. Asian J. Plant Sci., 4: 652-659.
CrossRefDirect Link - Agbagwa, I.O. and B.E. Okoli, 2005. Chromosome number and cytomorphological characterization of a polyploid Abrus. Afr. J. Biotechnol., 4: 607-610.
Direct Link - Agbagwa, I.O. and B.E. Okoli, 2006. Leaf epidermal micromorphology in the systematics of Abrus (Papilionaceae) in parts of tropical West Africa. Asian J. Plant Sci., 5: 41-49.
CrossRefDirect Link - Chen, X. and J. Wen, 2005. Leaf anatomical acclimation of six tree species to low soil water content. Int. J. Bot., 1: 212-218.
CrossRefDirect Link - Hoareau, L. and E.J. da Silva, 1999. Medicinal plants: A re-emerging health aid. Electron. J. Biotechnol., 2: 56-70.
Direct Link