
Salim H. Al-Rawahy
Department of Biology, College of Science, Sultan Qaboos University, P.O. Box 36, Alkhod 123, Sultanate of Oman
Haitham Al-Amri
Department of Biology, College of Science, Sultan Qaboos University, P.O. Box 36, Alkhod 123, Sultanate of Oman
Abdulrahman Al-Hinai
Department of Biology, College of Science, Sultan Qaboos University, P.O. Box 36, Alkhod 123, Sultanate of Oman
Neelam Sherwani
Department of Biology, College of Science, Sultan Qaboos University, P.O. Box 36, Alkhod 123, Sultanate of Oman
Asian Journal of Plant Sciences
Year: 2007 | Volume: 6 | Issue: 4 | Page No.: 673-677







ABSTRACT
The present study aimed to investigate the sensitivity of an indigenous leguminous plant species of Oman, Prosopis cineraria (ghaff tree), to SO2 pollutant. Plants were exposed to 0, 25, 50, 100 and 150 ppb SO2 for 30 min daily for the period of ten weeks, under light and dark conditions. The formation of marginal necrotic areas on leaflets was seen as the first symptom of SO2 injury in P. cineraria plants. Leaf senescence was highly significant (p<0.01) in plants exposed to SO2 in light conditions and significant (p<0.05) in plants exposed to SO2 in dark conditions compared with control plants. There was significant (p<0.05) decrease in Relative Growth Rate per week in plants exposed to SO2 in both light and dark treatments compared with control plants, but more pronounced reduction in light conditions. Stomatal conductance was significantly (p<0.01) reduced after SO2 exposure in both light and dark treatments. These results became the first record of Oman indigenous plant species showing confirmed injuries as a result of exposure of SO2 concentrations.
PDF Abstract XML References Citation
How to cite this article
Salim H. Al-Rawahy, Haitham Al-Amri, Abdulrahman Al-Hinai and Neelam Sherwani, 2007. Growth and Stomatal Conductance of Prosopis cineraria (Ghaff Tree) Exposed to Sulphur Dioxide. Asian Journal of Plant Sciences, 6: 673-677.
DOI: 10.3923/ajps.2007.673.677
URL: https://scialert.net/abstract/?doi=ajps.2007.673.677
DOI: 10.3923/ajps.2007.673.677
URL: https://scialert.net/abstract/?doi=ajps.2007.673.677
INTRODUCTION
Sulphur dioxide (SO2) is an industrial air pollutant associated with fossil fuel combustion and refining. It is one of the several pollutants released in automobile exhaust fumes, industrial smokestacks and metal smelting effluent. Together, natural and anthropogenic global SO2 sources emit an estimated 194 million tones annually, of which about 83% is anthropogenic from fossil fuel combustion (Botkin and Keller, 2005). Once emitted, SO2 is deposited onto surfaces by diffusion at various rates according to meteorological conditions, or may undergo a number of chemical reactions before dry or wet deposition. Among S gases, SO2 is considered to be the most important phytotoxic molecule (Jacobson, 2002) and its adverse effects on plants were recognized long before the effects of the other air pollutants (Winner et al., 1985). Recently, Swanepoel et al. (2007) reported that increased uptake of SO2 causes toxicity and reduced growth and productivity in plants due to accumulation of sulphite and sulphate within cells.
In some areas of Oman, especially in the vicinity of industrial areas, the visible foliar injury was observed in some indigenous plant species, e.g., Prosopis cineraria. Plants growing within these areas are exposed to greater SO2 concentrations as well as other pollutants, such as ozone (O3) and nitrogen dioxide (NO2). Hence there is considerable concern about the interactive and synergistic effects of exposure to mixtures of SO2 and other pollutants (Ashmoor, 2002). However, it is important as a first step that the effects of each pollutant be understood independently, that can help identify similarities in the ways in which environmental pollutants effect vegetation and contribute to our general understanding of plant responses to combination of pollutants. To evaluate the impact of contamination in an ecosystem, it is necessary to first establish the background level of the contaminants. The background level may be interpreted as a natural level, that is the average conditions of an area where there may be human activity, but which is in a good state of conservation (Conti and Cecchetti, 2001). From continuous air quality monitoring in Oman during the past 5 years (unpublished data), the background concentration of SO2 in rural area has been recorded as 11 ppb, while the average background concentration in cities is 22 ppb. Frequently (average once per week), however, the SO2 concentration in Muscat (capital) area exceeds 50 ppm and occasionally (average once per two months) exceeds 100 ppb. Once the background level had been established, the contamination factor may be used to evaluate the state of conservation or degradation (Conti and Cecchetti, 2001). The majority of past researches on the effects of SO2 on plants were focused on plants of economic value. Recent concern about the natural environment, however, has focused more interest on the ecological value of ecosystems. This study aimed to investigate the effects of SO2 exposure with special attention to growth and stomatal conductance in indigenous leguminous plant species of Oman, Prosospis cineraria. The species is an excellent multi-purpose tree for local people, particularly in providing fodder, fuel-wood and shade protection, as well as creating microenvironment that supports various wildlife (Brown, 1991).
MATERIALS AND METHODS
Seeds of P. cineraria (L.) Druce collected from wild trees when they became available in the summer of 2004 and 2005. Seeds were germinated in pots containing uniform soil compost. Seedlings were grown under growth room conditions (14 h photoperiod, flux density of 110 μmoles m-2 sec-2 at 28±2°C and 70±5% RH) and irrigated daily with tap water for 4 months. Fifty plants were then selected randomly and exposed to 0 (control) 50, 100 and 150 ppb SO2 gas (from MEGS Specialty Gases, Inc.) for 30 min daily for 10 weeks in a 150x100x65 cm perforated glass chamber. SO2 flow rate was manually adjusted using SUPERIORTM Gas Sulphonator and the concentration within the chamber was measured using a Tetra Crowcon SO2 gas detector. The experiment was repeated under light and dark conditions. In dark treatment light was switched off 30 min before the exposure to SO2 and switched on immediately after the exposure. Stomatal conductance of six plants per treatment selected randomly was measured on the first day of the experiment starting 1 h before the exposure to SO2 and 5 h after the exposure (-60, 0, 60, 120 180, 240 and 300 min) by using LI-COR, LI-6200 Portable Photosynthesis System and LI-6250 Gas Analyser. In order to understand the stomatal behaviour of P. cineraria in field conditions, 10 wild plants were randomly selected at various locations between 23°20’N-57°45’E and 22°40’N-58°35’N. Stomatal conductance of each plant was measured at least 2 different days during the month of August 2005 at interval of 1 h between 6.00 and 18.00 h. Experimental plants were observed for any foliar injuries and these were assessed in percentages (No. of injured leaves/total No. of leaves x 100). Observation of minute necrotic spots was made using the light microscope. At the end of the experiment, plants were harvested and the Relative Growth Rate (RGR) per week of each plant was calculated:
RGR/week = (ln (Dry Weight)-ln(Σ Initial Dry Weight))/10.
Initial dry weight is the average dry weight of 25 plants selected randomly harvested just prior to the first exposure of SO2 to above experimental plants. Data were analysed using a factorial design Analysis of variance (ANOVA) using SPSS package and presented as means±standard error.
RESULTS
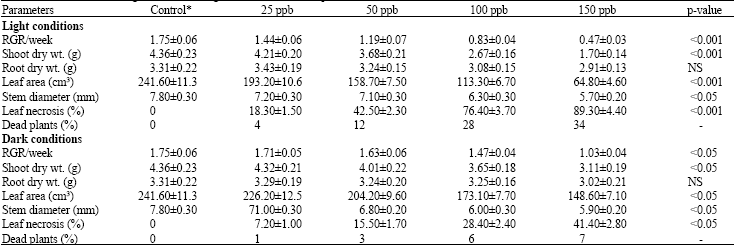
The formation of marginal necrotic areas on leaflets was seen as the first symptom of SO2 injury ( Table 1). These marginal necrotic areas started as dark green color and eventually became dry and changed to brown color. At the end of the experiment, 89 and 41% of the total number of leaves observed had SO2 damage symptoms under light treatment and dark treatment, respectively.
The difference in the number of leaves injured was highly significant (p<0.01) between different SO2 concentrations under light conditions and significant (p<0.05) under dark conditions. Minute necrotic spots around or near stomata were observed under the light microscope, which were significantly more numerous in plants exposed to SO2 under light conditions. There were no observed foliar injuries in control plants.
| Table 1: | The effects of fumigated SO2 on the growth of P. cineraria plants |
 | |
| *Only one set of control treatment, grown under growth room conditions, NS: Non significant | |
| Table 2: | Stomata conductance (mmol m-2 sec-1) of P. cineraria as affected by exposure of different concentration of fumigated SO2 for 30 min. Stomata conductance measurements of control plants under light condition were made only once |
 | |
| *60 min before SO2 exposure, +Measurements start immediately after SO2 exposure and last for 30 min, NS: Non significant | |
| Table 3: | Stomata conductance of 10 wild P. cineraria plants measured during the month of August 2005 at interval of 2 h |
 | |
| *n = 2 | |
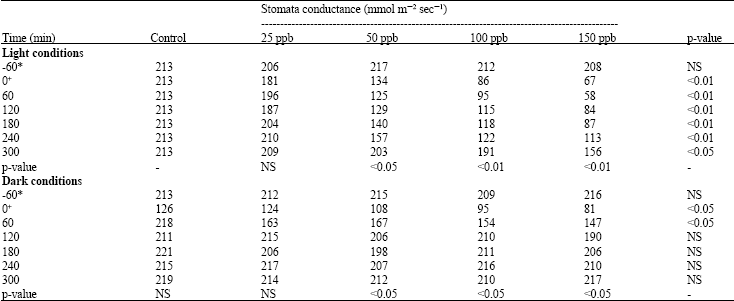
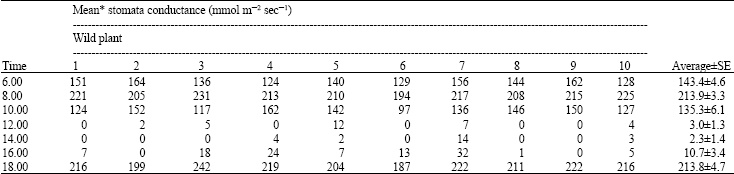
Leaf senescence was mostly observed in plants exposed to SO2 under light conditions. Compared to the control plants, there was a highly significant (p<0.01) decrease in RGR/week in plants exposed to SO2 under light treatment and a significant (p<0.05) decrease under the dark treatment compared with control plants. Stem diameter was significantly (p<0.05) reduced in exposed plants under both light and dark conditions. Stomatal conductance was substantially reduced after SO2 exposure under light conditions compared to SO2 exposure under dark conditions ( Table 2). These responses were transient in dark treatment and maintained for several hours after SO2 exposure under light treatment. Stomatal conductance in wild plants was greater during the early and late hours of the day ( Table 3).
DISCUSSION
In this study, the symptoms of SO2 injury were first seen on the leaves of P. cineraria. Almost 75% of the overall population exposed to SO2 has showed visible symptoms and incidence of injury was greater for plants exposed to SO2 under light condition. Descriptions of visible injuries and susceptibility of many plant species to SO2 have been reported by a number of investigators (Bell and Mudd, 1976; Ayazloo and Bell, 1981; Keller, 1981; Black, 1982; Krupa, 1996; Legge and Krupa, 2002; Moraes et al., 2002; Raziuddin et al., 1999). Chlorosis and marginal necrotic areas on leaves are the prominent phenomenon of SO2 phytotoxicity and are derived from the breakdown of photosynthetic pigments in mesophyll tissues (Garsed, 1985). The necrotic areas in this study ranged in color from dark green to reddish-brown to brown. This is supported by SO2-induced damage reported by Legge and Krupa (2002). Dry deposition of SO2 involves the transfer of SO2 from the air stream to the canopy (Rennenberg and Polle, 1994). Once in the canopy, SO2 may penetrate the boundary layer by a diffusion process. SO2 molecules that move through boundary layer of P. cineraria canopy will most probably enter the leaves through stomatal openings. Garsed (1985) reported that the pathway of SO2 to unwetted foliage of Vicia faba is largely through open stomata and less than 10% of the total flux was actually accounted for by adsorption on the cuticle. The absorption to the surface of the leaves characterized by a thick cuticle layer could be very much reduced. The diffusion flux of SO2 molecules to the site of assimilation or damage in the leaf mesophyll is determined by the concentration gradient and leaf diffusion resistance (Heldt, 1996). Following the absorption of SO2 through the stomatal pores the gas is dissolved in mesophyll spaces near the stomata to form hydrated SO2 (SO2.H2O), which act as a strong acid, dissociating to HSO3− and SO3− in proportions determined by the pH of the water films bounding the mesophyll epidermal apoplast (Legge and Krupa, 2002). Peiser and Yang (1985) reported that SO2 is rapidly hydrated, forming bisulfite and sulphite. These byproducts have been implicated in SO2 toxicity and if the uptake exceeds the capacity of cells to detoxify sulfites, minute necrotic spots are formed near or around stomata.
In this study the foliar SO2 injury symptoms were mostly seen on leaves at a full stage of development. Similar SO2 injury symptoms were observed in other plant species including Alsike clover (Trifolium hybridum) and prickly rose (Rosa acicularis) (Legge and Krupa, 2002). Various studies showed that there is a strong correlation between leaf senescence in plants and SO2 exposure (Heldt, 1996).
The results presented here showed highly significant difference in leaf senescence in plants exposed to SO2 under light conditions and significant in plants exposed to SO2 in dark conditions compared with control plants. If the rate of leaf senescence becomes faster than the rate at which new leaves grow, the photosynthetic leaves will decrease and this in turn will reduce net assimilation rates and relative growth rates. SO2 can also affect photosynthesis by altering stomatal conductance (Marshall, 2002) or by changing the metabolic capacity of mesophyll cells (Winner et al., 1985). Mansfield and Pearson (1996) reported that short-term exposure to SO2, particularly at concentration <50 ppb, often causes wider stomatal opening, while long-term exposure with higher concentrations usually causes partial stomatal closure. Present results with P. cineraria showed that stomatal closure can occur within minutes after the SO2 exposure of 50, 100 and 150 ppm under light treatment and this response is maintained for a few hours. In field measurements of 10 wild P. cineraria plants, it was found that stomatal conductance was greater during early and late hours of the photoperiod. The morning hour peak and late evening peak of SO2 levels during most days in the vicinity of industrial areas and urban areas may directly alter natural stomatal behavior in this plant species. Photosynthetic decline in P. cineraria in the presence of SO2 pollution could therefore be directly related to reduced stomatal conductance. Moreover, the proportion of photosynthetic inhibition due to non-stomatal factors, including physiological and biochemical damage, increase as greater quantities of SO2 are absorbed into the leaf (Krupa, 1996). The advantage of stomatal closure is the reduction or alteration in the quantity of SO2 that enters the plant and arrives at metabolic sites. The disadvantage, however, is the depression in photosynthetic carbon dioxide uptake (Raziuddin et al., 1999). The level of leaf physiological responses of decreased photosynthetic capacity that lead to altered carbon allocation pattern may ultimately influence growth characteristics such as shoot or root growth (Al-Rawahy, 2000). Decreased growth may affect plant’s ability to acquire essential resources from the environment (Novak et al., 2003).
In response to many stresses, shoots are affected more than roots (Al-Rawahy, 2000). This was also the case of P. cineraria seedlings, where there was no significant reduction in root dry weight after SO2 exposure compared with control seedlings in both light and dark treatments. Cheesman (1993) suggested that increase root/shoot ratio in stressed plants help to reduce the demand for photosynthetic products to shoot while maintain the root size so the absorption of water and mineral is not affected. However, the cost for this change is the reduced ability to supply products of photosynthesis to the growing apices (Al-Rawahy et al., 2003). In the long term exposure to stress the growth is likely to be strongly reduced even in roots.
The current study was successful in validating SO2-induced foliar injury and reduction in stomatal conductance in P. cineraria plant species. These results became the first record of Oman indigenous plant species showing confirmed injuries as a result of exposure of SO2 concentrations. Several genera and species need to be investigated in order to establish which are the more sensitive species that may serve as bioindicators of SO2 pollutants in the field. Use of native plant species as bioindicators would have a higher significance for the characterization of the air pollution impact on the ecosystem than those from studies with exotic plant species (Moraes et al., 2002).
ACKNOWLEDGMENT
The Authors would like to thank Sultan Qaboos University for supporting research and providing facilities. The Ministry of Regional Municipalities, Environment and Water Resources is very much thanked for various support and help throughout the project. Dr. M. Kokkin, Department of Biology, SQU, is thanked for reading the manuscript.
REFERENCES
- Al-Rawahy, S.H., K.S. Al-Dhafri and S.S. Al-Bahlany, 2003. Germination, growth and drought resistance of native and alien plant species of the genus Prosopis in the sultanate of oman. Asian J. Plant Sci., 2: 1020-1023.
CrossRefDirect Link - Ayazloo, M. and J.N.B. Bell, 1981. Studies on the tolerance to sulphur dioxide of grass populations in polluted areas. 1. Identification of tolerant populations Dactylis glomerata, Festuca rubra, Holcus lanatus. New Phytol., 88: 203-222.
PubMed - Cheesman, J.M., 1993. Plant growth modeling without integrating mechanisms. Plant Cell Environ., 16: 137-147.
CrossRef - Conti, M.E. and G. Cecchetti, 2001. Biological monitoring: Lichens as bioindicators of air pollution assessment: A review. Environ. Pollut., 114: 471-492.
CrossRefDirect Link - Keller, T., 1981. Winter uptake of airborne SO2 by shoots of deciduous species. Environ. Pollut., 26: 313-317.
CrossRef - Moraes, R.M., A. Klumpp, C.M. Furlan, G. Klumpp, M. Domingos, M.C.S. Rinaldi and I.F. Modesto, 2002. Tropical fruit trees as bioindicators of industrial air pollution in southeast Brazil. Environ. Int., 28: 367-374.
CrossRefDirect Link - Novak, K., J.M. Skelly, M. Schaub, N. Krauchi, C. Hug, W. Landolt and P. Bleuler, 2003. Ozone air pollution and foliar injury development on native plants of Switzerland. Environ. Pollut., 125: 41-52.
CrossRefDirect Link - Swanepoel, J.W., G.H.J. Kruger and P.D.R. Heerden, 2007. Effects of sulphur dioxide on photosynthesis in the succulent Augea capensis Thunb. J. Arid Environ., 70: 208-221.
CrossRefDirect Link