
S.A. Ogunbayo
Africa Rice Center, 01 BP 2031, Cotonou, Benin Republic
D.K. Ojo
Department of Plant Breeding and Seed Technology, University of Agriculture, Abeokuta, PMB 2240, Nigeria
A.R. Popoola
Department of Plant Breeding and Seed Technology, University of Agriculture, Abeokuta, PMB 2240, Nigeria
O.J. Ariyo
Department of Plant Breeding and Seed Technology, University of Agriculture, Abeokuta, PMB 2240, Nigeria
M. Sie
Africa Rice Center, 01 BP 2031, Cotonou, Benin Republic
K.A. Sanni
Africa Rice Center, 01 BP 2031, Cotonou, Benin Republic
F.E. Nwilene
Africa Rice Center, 01 BP 2031, Cotonou, Benin Republic
E.A. Somado
Africa Rice Center, 01 BP 2031, Cotonou, Benin Republic
R.G. Guei
FAO Viale delle Terme di Caracalla, 00100 Rome, Italy
D.D. Tia
Africa Rice Center, 01 BP 2031, Cotonou, Benin Republic
O.O. Oyelakin
International Institute of Tropical Agriculture (IITA), PMB 5320, Ibadan, Nigeria
A. Shittu
Africa Rice Center, 01 BP 2031, Cotonou, Benin Republic
Asian Journal of Plant Sciences
Year: 2007 | Volume: 6 | Issue: 4 | Page No.: 653-666







ABSTRACT
Morphological and molecular characterization of ninety-six landrace rice (Oryza sativa L.) accessions including six checks collected from four regions (North-West, North, West and Central-West) of Cote d’Ivoire were assessed using 14 agro-botanical traits and 10 Randomly Amplified DNA Polymorphisms (RAPDs) primers, respectively. Accessions were evaluated in a field experiment in an augmented experimental design with three replicates. The aim of the research was to study variations and to select lines that could be used as potential parents in future breeding programs. A principal components plot and a dendrogram based on distance between genotype cluster groups for mean values of the morphological variables were used to group the accessions. Genetic relatedness among accessions based on RAPD molecular marker was also presented in form of a dendrogram generated by clustering analysis using the Unweighted Pair Group Method with Arithmetic mean (UPGMA). The relative effectiveness of the RAPD markers compared to botanical descriptors in assessing diversity among the accessions was investigated. Clustering analysis technique using NTSYS classified the 96 landrace accessions into 8 morphological groups whereas PCA re-ordered the accessions into three broad groups that had within cluster similarities and inter-cluster differences in morphological variations. Reaction products (bands) of the RAPD analysis were highly polymorphic, more discriminatory and informative as they were able to differentiate more pairs of accessions than the botanical descriptors. Apart from checks, highest grain yield (2316 g/plot) was observed for accessions 46 (DNN 184) with an average of 12 filled tillers, plant height of 136 cm and medium maturity date of 136 days. It was observed that number of total and filled tillers per se was not a function of yield but rather, these traits were significantly associated with plant height and maturity date. Although, landrace rice accessions in Côte d’Ivoire is associated with relatively narrow genetic base, positive heterosis could be promoted if any of the Gagnoa (GGA) accessions from Central West of the country is used in a future hybridization program with Danane (DNN) accessions from west because of genetic distance between members of the two groups.
PDF Abstract XML References Citation
How to cite this article
S.A. Ogunbayo, D.K. Ojo, A.R. Popoola, O.J. Ariyo, M. Sie, K.A. Sanni, F.E. Nwilene, E.A. Somado, R.G. Guei, D.D. Tia, O.O. Oyelakin and A. Shittu, 2007. Genetic Comparisons of Landrace Rice Accessions by Morphological and RAPDs Techniques. Asian Journal of Plant Sciences, 6: 653-666.
DOI: 10.3923/ajps.2007.653.666
URL: https://scialert.net/abstract/?doi=ajps.2007.653.666
DOI: 10.3923/ajps.2007.653.666
URL: https://scialert.net/abstract/?doi=ajps.2007.653.666
INTRODUCTION
Rice (Oryza sativa L.) is one of the most important crops that provide food for about half of the world population, particularly in Asia, Africa and Latin America where the demand for rice is a top priority (Sasaki, 1999, 2002). Its adoption as a principal staple food is increasing in Africa whereas self sufficiency in rice production is declining as demand increases. Hence, there is an urgent need to increase and improve the production of rice in Africa in order to meet up with the high demand (Ogunbayo et al., 2005). The need for expansion of rice cultivation does not only depend on cultural practices and management, it also depends on the suitability of rice varieties, which must be drawn from existing germplasm that has been collected and conserved by genetic resources centers (Ng et al., 1988).
Evaluation and characterization of landrace rice should form an important constituent of collection efforts because of their enormous in-built genetic diversity due to several generations of growing and selection by breeders and farmers. Landraces also constitute a good source of unique genes for stress tolerance, high yield stability, adaptability to the environments and genetic dynamics (Frankel et al., 1995; Guei and Traore, 2001). However, the utilization of these rice genetic resources had been limited to only adaptable genotypes (Caldo et al., 1996). Thus, a successful breeding programme will depend on the genetic diversity of a crop for achieving the goals of improving the crop and producing high yielding and better resistant varieties (Padulosi, 1993). A number of landraces are still cultivated locally but most of them are being rapidly replaced by improved cultivars due to increasing narrowing of genetic base. The adoption of new varieties means that the area planted to landraces are gradually disappearing (Guei, 2000). Thus, reduced genetic variability underscores the need to collect landraces for ex situ conservation and to characterize them for future rice breeding programs at morphological and molecular levels because the evaluation of phenotypic diversity usually reveals important traits of interest to plant breeders (Singh, 1989).
Ogunbayo et al. (2005) carried out a phylogenetic evaluation of forty rice accessions using morphological and molecular techniques in which within cluster similarities and between cluster morphological differences were observed. Landrace rice differ from improved cultivars in adaptation to soil type, sowing and ripening periods and yield stability particularly, in regions where seasons are unpredictable (Frankel et al., 1995). However, they constitute a good source of unique genes for stress tolerance (Guei and Traore, 2001) and they are genetically dynamic. Despite these positive attributes, little efforts have been made to characterize and evaluate landrace rice accessions of West Africa origin. The objectives of this study were to investigate the morphological and molecular organizations of the existing diversity in ninety-six landrace rice accessions and to compare genetic relatedness among these accessions.
MATERIALS AND METHODS
Evaluation of morphological traits: The germplasm used in this study consisted of ninety-six landrace accessions including ten improved varieties in which six were checks. The materials were collected by the Genetic Resources Unit of the Africa Rice Center (WARDA) from four different localities Gagnoa, Danane, Touba and Boundiali and their respective surrounding villages, in the West-central, Western, North-Western and Northern parts of Côte d’Ivoire, West Africa ( Fig. 1). The collections have since been maintained ex situ in the gene bank of Africa Rice Center (WARDA) at M’bé, Côte d’Ivoire. For the purpose of easy identification and retrieval, accession number comprising of three letters (to represent the site), plus a serial number was assigned to each collection. Thus, landrace collected from Danane bore the prefix DNN Gagnoa GGN while those from Touba and Boundiali bore the prefixes TBA and BDL, respectively.
The experimental study was conducted under irrigated lowland conditions during the year 2004 and 2005 wet seasons at International Institute of Tropical Agriculture (IITA), Ibadan, Nigeria. The augmented experimental design introduced by Federer (1956, 1961, 1991) was used for the experiment. An Augmented Experimental Design (AED) is usually useful for testing a large number of genotypes in early generations when valid statistical analyses are needed particularly when seed supplies are too limited to permit replication. The basic concept of augmented design construction is to establish a standard replicated design using checks for which sufficient seeds are available. Each replicate forms a complete block, incomplete block, or cell, depending on the standard design. Additional unassigned plots are created within each replicate and un-replicated entries for which there are insufficient seeds. Entries are then assigned to these plots in the form of an incomplete block design. The seeds were sown in the nursery bed before they were transplanted at 21 day old with a spacing of 20x20 cm. One seedling was transplanted per hill and the inter-plot spacing was 40 cm. A plot size of 1.2x5 m with 6 rows was used for each accession in the field.
Recommended cultural practices for the evaluation included fertilizer NPK (15-15-15) as basal application at a rate of 150 kg ha-1 during land preparation and urea was applied at the rate of 50 kg ha-1 as top-dressing first at tillering and a second time at booting. Morphological data were collected for fourteen quantitative characters at appropriate growth stage of rice plant following the descriptor for Rice Oryza sativa L. (IRRI, 1980). The characters that were evaluated are plant height, leaf length, leaf width, days to 50% heading, days to maturity, total No. of tillers at heading, No. of fertile tillers, panicle shattering, tiller diameters, panicle length, grain length, grain width, 100 grain weight and yield per plot. The data collected on 14 agro-botanical traits from the rice accessions were subjected to statistical analysis using SAS/PC version 9.1 (SAS, 1999) and NTSYS pc 2.0 (Rohlf, 1993). Principal components grouping of the traits was employed to examine the percentage contribution of each trait to total genetic variation. Cluster analysis based on similarity matrices was also employed on agro-botanical data using the un-weighted pair group method with arithmetic mean (UPGMA) to obtain a dendrogram.
PHENOL REACTION
As mentioned by Oka (1958), the phenol reaction is a good criterion to discriminate between Oryza sativa sub-species, Indica and Japonica and it is equally effective with African rice (Kochko de, 1987). All the varieties were tested for their response to the phenol reaction and the grains were soaked in a 2% phenol solution for 48 h. The change in color of the hull was compared with that of grains soaked in distilled water (control) for the same period and comparison of grains, based on the change in color of the hull were indicated as (+) and (-) in Table 5.
RAPD analysis: Total DNA was isolated from the leaves of 7 day-old seedlings, grown in the green house, according to Dellaporta et al. (1983). Gene-based RAPD analysis was performed on ninety-six rice populations from the species O. sativa and O. glaberrima. Purified DNA was quantified by spectrophotometry and by ethidium bromide coloration after electrophoresis. Ten RAPD primers were used to generate markers as described by Tao et al. (1993). Each amplification was performed in a reaction volume of 25 μL containing 10 mM Tris-HCl pH 9.0, 50 mM KCl, 1.5 mM MgCl2, 0.2 mg mL-1 gelatin, Triton x100 0.1%, 0.1 mM of each dATP, dCTP, dGTP and dTTP (Promega), 10 ng of random primer, 50 ng of genomic DNA and 2 units of Taq polymerase. Amplification was carried out in a Mpi thermocycler as follows: 1 cycle of 3 min at 94°C; 44 cycles of 20 sec at 94°C; 40 sec at 37°C and 1 min at 72°C; 1 cycle of 7 mins at 72°C. Amplification products were then analysed for polymorphism after electrophoresis in 1.2% agarose gels in 0.5X TBE buffer and stained with ethidium bromide. Pairwise comparison of genotypes, based on the presence (1) or absence (0) of unique and shared polymorphic products was used to generate similarity coefficients using statistical software package NTSYS-pc 2.0 (Rohlf, 1993) and Jaccard coefficient of similarity (Jaccard, 1908). Genetic diversity (bp) was computed from the binary data for all pairwise combinations of accessions according to the Nei diversity index (Nei, 1972). The similarity coefficient was used to construct a dendrogram by the unweighted pair group method with arithmetic average (UPGMA) according to Sneath et al. (1973), Swofford et al. (1990) and Rohlf (1993).
RESULTS
Significant block effects were observed for leaf width, grain length and 100 grain weight whereas block effects were non-significant for the other traits meaning that blocking was not important for the eleven traits that showed non-significant block effect ( Table 1). The six checks in each replicate as well as the accessions differed significantly with respect to all 14 traits.
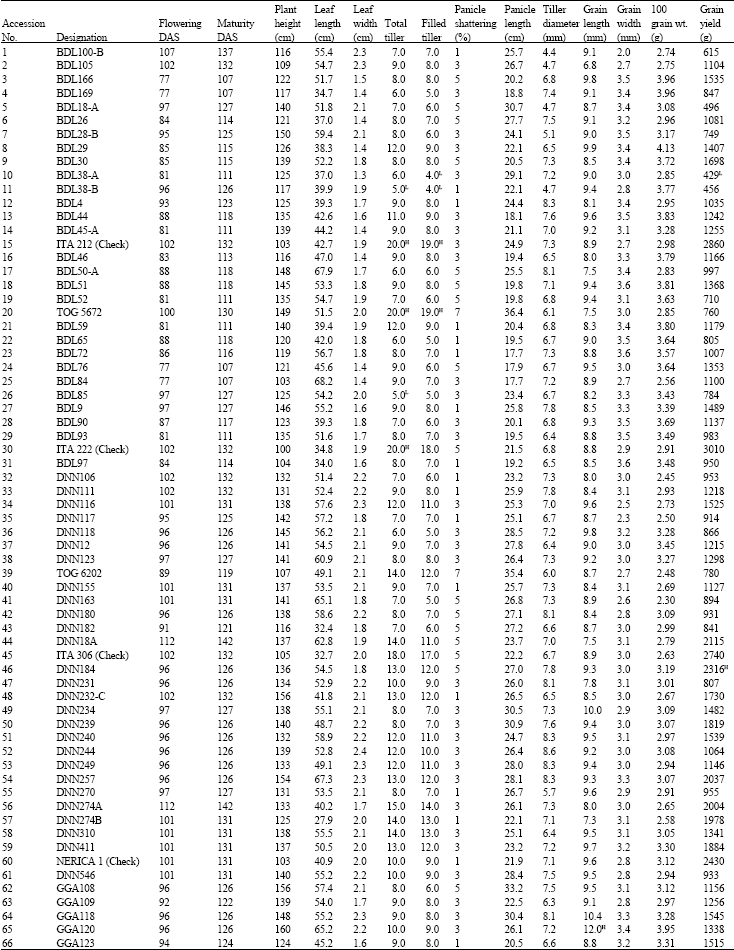
Accessions 92 (CG 14) recorded the earliest flowering date (75 days), earliest maturity date (105 days) as well as longest panicle length (38.7 cm) ( Table 2). CG 14 also had a yield of 1985 g per plot. However, accessions 68 (GGA 136) recorded the longest flowering date (113 days), longest maturity date (143 days), shorter panicle length of 32.2 cm and a yield of 1965 g per plot that was not statistically different from that of CG 14. Accession 93 (TBA 1) with shortest panicle length of 15.6 cm had a yield of 806 g per plot and 26 (BDL 85) had the least number of total tillers (5) whereas accession 20 had the highest (20). Also, accessions 10 and 11 had the least No. of filled tillers (4) whereas accessions 15, 20 recorded the highest No. of filled tillers, 19 in each case. Apart from the checks, highest grain yield per plot was observed for accession 46 (DNN 184) that had an average of 12 filled tillers, plant height of 136 cm and medium maturity date of 126 days. However, accession 10 (BDL 38-A) with an average No. of total and filled tillers of 6 and 4, respectively, recorded the least (429 g) grain yield per plot. Interestingly, accession 20 (TOG 5672) that had the highest No. of total (20) and filled (19) tillers recorded a yield of 760 g per plot probably because the entry was too tall (149 cm) and late maturing (130 days) compared with the highest yielder (DNN 184). It should also be noted that most of the GGA accessions had average plant height and thus, average yield per plot.
The correlation matrix showed that grain yield was positively and significantly associated with maturity date, total tiller, filled tiller, panicle length and grain length ( Table 3). However, grain yield had negative but significant association with grain width, leaf length, leaf width, panicle shattering, tiller diameter and 100 grain weight.
| Table 1: | Mean squares from analysis of variance for augmented randomized complete block design for fourteen traits measured in ninety six rice accessions |
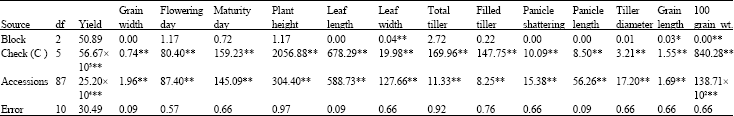 | |
| *, ** Significant at 5 and 1% probability levels, respectively | |
| Table 2: | Means of fourteen characters measured in ninety-six rice accessions |
 | |
| Table 2: | Continued |
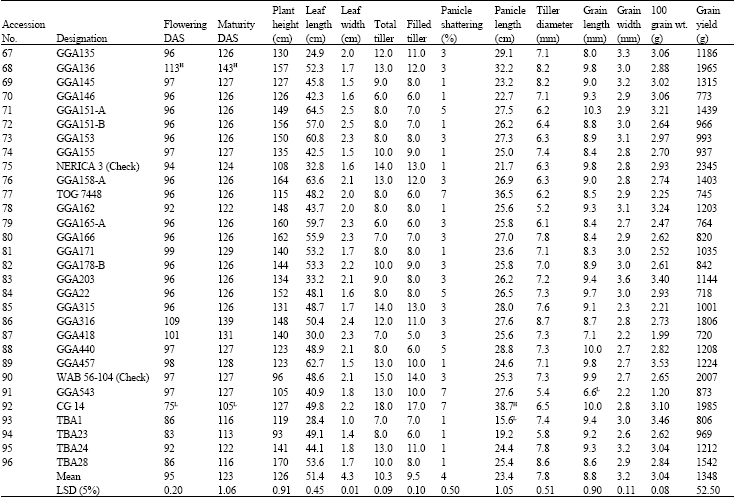 | |
| H: High, L: Low | |
| Table 3: | Correlation coefficients of fourteen traits used in characterizing ninety-six rice accessions |
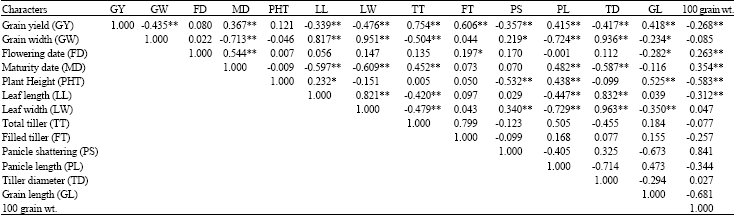 | |
| *, **, Significant at 5% and 1% probability levels, respectively | |
Grain width was positively and significantly correlated to leaf length, leaf width, panicle shattering and tiller diameter. While flowering day were also positively and significantly correlated to maturity days, filled tiller, grain weight and maturity days were positively and significantly correlated to total tiller, panicle length and grain weight.
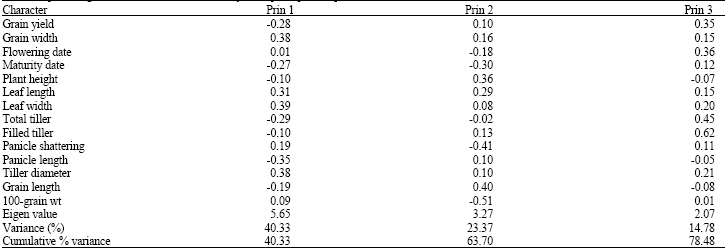
The three principal components accounted for about 78.48% of total variance with the first principal component taking 40.33% ( Table 4). The relative discriminating power of the principal axes as indicated by the eigen values was highest (5.65) for axis 1 and lowest (2.07) for axis 3.
The first principal component that accounted for the highest proportion (40.33%) of total variation was mostly correlated with leaf width, tiller diameter, grain width, leaf length, panicle length, total tiller and grain yield.
| Table 4: | Principal components analysis showing the contribution (factor scores) of each character among the ninety-six rice accessions, eigen values and percentage total variance accounted for by three principal components |
 | |
| Table 5: | Phenol reaction that characterized the ninety-six rice accessions into Indica (Lowland) and japonica (Upland) sub-species |
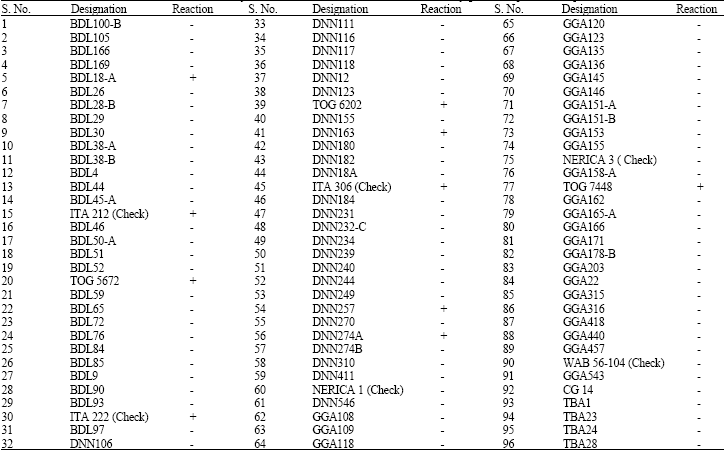 | |
| + = Indica (Lowland), - = Japonica (Upland) | |
Characters that were mostly correlated with the second principal component were grain length, plant height, leaf length, 100-grain weight, maturity date and panicle shattering. The third principal component was dominated by traits such as filled tiller, total tiller, flowering date, grain yield and leaf width. Out of ninety-six accessions only ten were reacted positively to phenol and thus classified as Indica while other were Japonica ( Table 5).
The ten operon primers generated a total of 108 RAPD bands all of which were polymorphic across accessions because they were able to differentiate at least any two of the ninety-six rice accessions at a time ( Table 6). The number of bands per primer varied from eight to fourteen with an average of 10.8. Primers were able to produce fragments that varied from 250-3000 bp in size. Residual heterogeneity within lines is suspected because all the RAPD primers were able to amplify more than one band per genotype.
| Table 6: | Nucleotide sequence of selected primers with the number of amplified products and fragment size range (bp) |
 | |
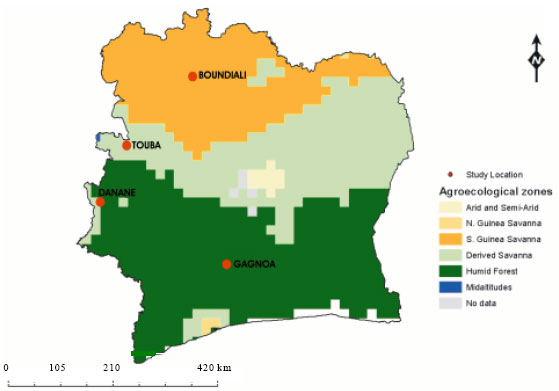 | |
| Fig. 1: | Map of Côte d’lvoire showing collection location |
Accessions within clusters 1 and 7 were associated with about 20% similarity, those within clusters 5 and 6 had about 12.5% similarity whereas accessions within clusters 2, 3 and 4 were the most similar (about 6.3% similarity) ( Fig. 2). Thus, to promote additional morphological variability among the 96 landraces, hybridization between accessions in clusters 3 and 4 (with highest distance) in one hand and those in clusters 1 and 7 (with least distance) on the other hand would be the most desirable. The dendrogram in Fig. 4 as opposed to that in Fig. 2 tends to suggest that most landrace rice accessions were morphologically similar with an average of about 12.5% morphological diversity.
A plot of relationship between the 96 landrace rice accessions as shown by the first and second principal components axes (Prin 1 and Prin 2) is shown in Fig. 3. Four out five accessions within cluster group plus accessions 74 and 93 of cluster group 4 were distributed at the top right side of the first principal axis. Accessions 8, 81 and 90 in cluster group 4 plus accession 83 in group 1 were also distributed towards the lower left side of first principal axis. However, all other accessions except 1,2,3 in group 7 were centrally distributed more towards the second principal axis and were thus, probably, morphologically similar. The above observations suggest within and between cluster variations resulting from large morphological diversity between the ninety-six accessions that were evaluated.
Figure 5 presents the molecular dendrogram of genetic similarity among ninety-six rice accessions as revealed by UPGMA cluster analysis based on RAPDs marker. The dendrogram shows a clear separation of the accessions into fourteen groups and allow discrimination between all 96 accessions compared with eight for the morphological grouping. A close phylogenetic proximity between TBA 1, TBA 28 and TBA 24 in group 9, accessions 20 (Tog 5672) and 39 (Tog 6202) in group 12 was shown.
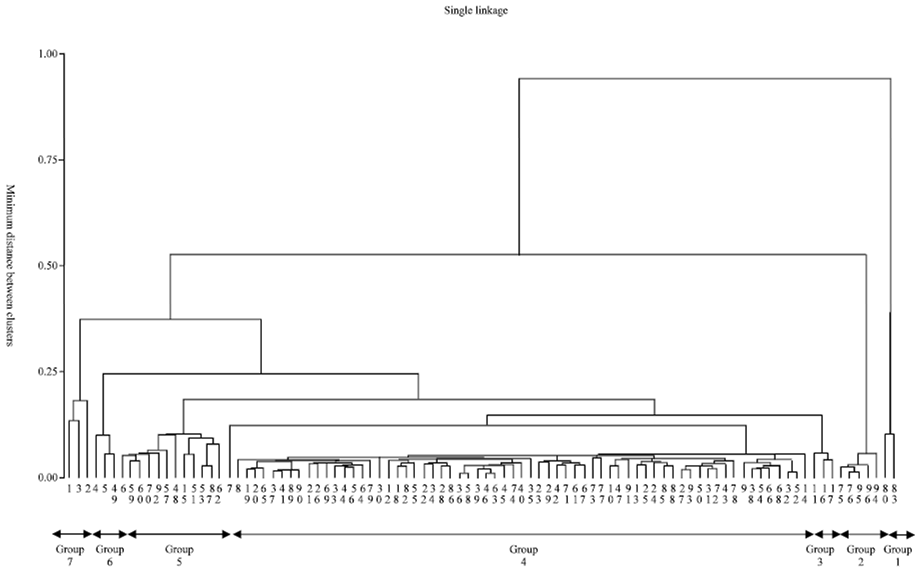 | |
| Fig. 2: | UPGMA-based morphological dendogram showing the minimum distance between clusters groups |
 | |
| Fig. 3: | Plot of prin 1 and 2 showing the relationship between clusters of ninety-six accessions (No. Represent accession numbers as in Table 3) |
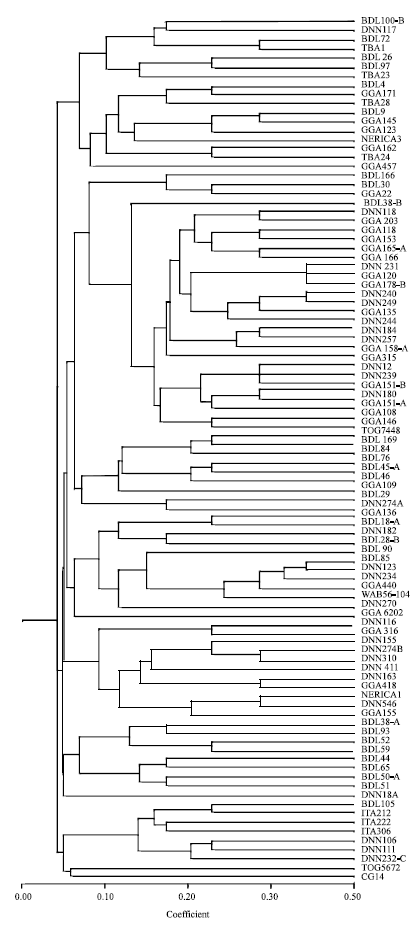 | |
| Fig. 4: | UPGMA-based morphological using the NTSYS-pc 2.0 showing the genetic similarity among ninety-six rice accessions |
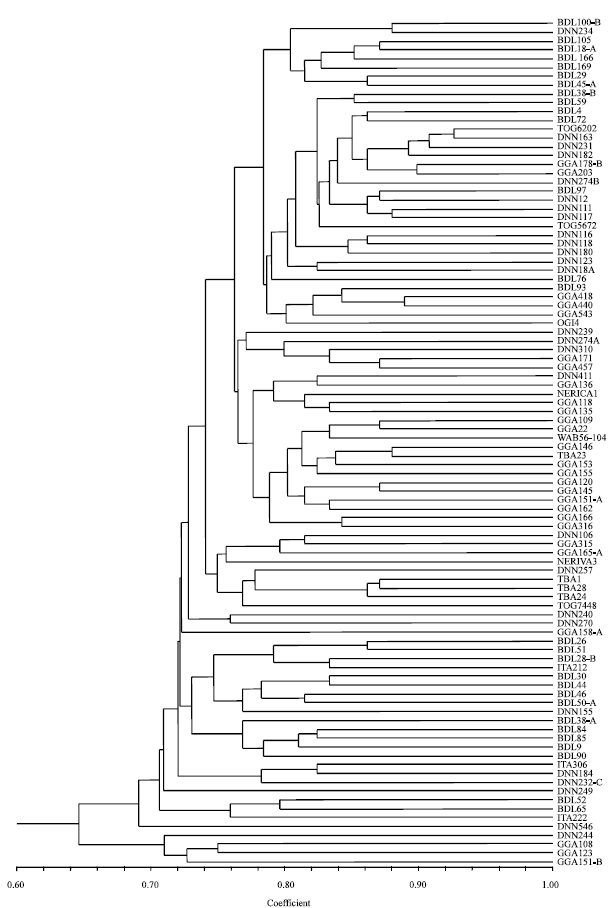 | |
| Fig. 5: | Molecular dendrogram showing the genetic similarity among ninety-six rice accessions revealed by gene-based RPADs |
These results confirm previous data based on morphological markers ( Table 2) that identified the five accessions as very similar in their vegetative apparatus and thus, very near from a taxonomical point of view. Pair of accessions 63 (GGA 109), 65 (GGA 120), 69 (GGA 145) and 84 (GGA 22) all in group 10 have very large (87.0%) similarity as they were from the same background. It was also observed that IITA accessions 15 (ITA 212), 30 (ITA 222) and 45 (ITA 306) were similar and had reasonable genetic similarity with accessions 7 (BDL28-B), 9 (BDL 30), 13 (BDL 44), 19 (BDL 52), 22 (BDL 65) and 46 (DNN 184).
Accessions 32 (DNN 106), 85 (GGA 315), 79 (GGA 165-A) and 75 (NERICA 3) in group 9 were genetically similar. Accessions 59 (DNN 411), 68 (GGA 136), 60 (NERICA 1) 64 (GGA 118) and 67 (GGA 135) in group 10 were equally genetically similar. However, the phenol test has indicated that all the above 9 accessions were japonica (upland) rice. Thus the RAPD marker has been able to confirm the phenol test. Heterosis can be promoted if any of the GGA accessions in group 1 can be used in a future hybridization program with accessions 39, 41, 47 in group 12 because of the genetic distance between members of the two groups.
DISCUSSION
Variations did exist among the ninety-six rice accessions with respect to the fourteen traits that were evaluated. Number of filled (fertile) tillers and total number of tillers on rice plants as well as maturity date were observed to greatly influence grain yield among the ninety-six rice accessions that were evaluated. On the other hand, flowering and plant height characteristics did not influence grain yield. Thus, it was not surprising that positive and significant associations were observed between yield and days to maturity (r = 0.367); total tiller (r = 0.754) and total filled tiller (r = 0.606). However, flowering date and plant height were insignificantly associated with grain yield because for instance accessions 68 (GGA 136) 92 (CG 14) that had the longest (113 cm) and shortest (75.0 cm) flowering dates recorded similarly grain yields.
The first principal component that accounted for the highest proportion (40.33%) of total variation was mostly correlated with leaf width, tiller diameter, grain width, leaf length, panicle length, total tiller and grain yield. Characters that were mostly correlated with the second principal component were grain length, plant height, leaf length, 100-grain weight, maturity date and panicle shattering. The third principal component was dominated by traits such as filled tiller, total tiller, flowering date, grain yield and leaf width.
According to Aliyu and Fawole (2000) cluster analysis has the singular efficacy and ability to identify crop accessions with highest level of similarity using the dendrogram. Evaluation of phenetic diversity within rice accessions using the cluster and PCA analyses in this study has provided seven and three clusters, respectively, with a lot of variations in morphological properties. Apart from the fact that the PCA was able to re-order accessions into three distinct clusters, the current study has shown that the use of morphological grouping could not provide a convincing discriminatory evidence in the classification of rice accessions. It only provided a sort of minimum distance between groups of accessions. Also, the morphological dendrogram generated from similarity or genetic distance matrices has provided an overall pattern of variation as well as the degree of relatedness among accessions.
However, due to variations in environmental conditions such as soil types and soil fertility levels (Steel, 1972), light, temperature and moisture regime (Summerfield and Huxley, 1973; Morakinyo and Ajibade, 1998), there is every tendency that different results are obtained using morphological grouping, particularly when experiments are repeated in time and/or space. Also, both the genetic make-up of seed, environment and field management practices have been reported to influence the morphology of a crop (Singh and Rachie, 1985). According to Virk et al. (2000), the use of morphological characters in the classification of germplasm, particularly in rice, has been met with difficulties because the technique is inefficient. Phenol reaction was able to classified the ninety-six rice accessions into sub-species group, indica (lowland) and japonica (upland). Out of ninety-six accessions only ten were reacted positively to phenol and thus classified as indica.
Observations above tend to emphasize the superiority of molecular similarity grouping over and above the morphological grouping. The fourteen major clusters of the RAPD dendrogram together with their internal groups have demonstrated the polymorphic nature of the ninety-six rice accessions used in the current study. The dendrogram obtained from the RAPD marker has revealed that the marker was more discriminatory, highly polymorphic and thus, more informative than the one obtained from morphological characterization because the marker was able to make use of ten OPERON primers to generate 108 RAPD bands across the ninety-six rice accessions. Consequently, some of the bands were polymorphic as each band was able to differentiate at least any two of the ninety-six rice genotypes. Thus, differentiation among rice genotypes was higher using the RAPD markers because these DNA markers are proven as efficient in the identification of phenotypic markers that are linked to agronomically important traits. Thus, such traits can be introgressed during the development of near isogeneic lines. The results are consistent with the morphological, allozyme and RFLP studies in sorghum (Dewet et al., 1978; Aldrich et al., 1992; Cui et al., 1995).
REFERENCES
- Aldrich, P.R., J. Doebley, K.F. Schertz and A. Stec, 1992. Patterns of allozyme variation in cultivated and wild Sorghum. Theor. Applieed Genet., 85: 451-460.
Direct Link - Caldo, R.A., L.S. Sebastian and J.E. Hernandez, 1996. Morphology-based genetic diversity analysis of ancestral lines of rice in Philippine rice cultivars. Philipp. J. Crop Sci., 21: 86-92.
Direct Link - Cui, Y.X., G.W. Xu, C.W. Magill, K.F. Schertz and G.E. Hart, 1995. RFLP-based assay of Sorghum bicolor (L.) Moench genetic diversity. Theor. Applied Genet., 90: 787-796.
Direct Link - De Wet, J.M.J., 1978. Systematics and evolution of sorghum sect. Sorghum (Gramineae). Am. J. Bot., 65: 477-484.
Direct Link - Dellaporta, S.L., J. Wood and J.B. Hicks, 1983. A plant DNA minipreparation: Version II. Plant Mol. Biol. Rep., 1: 19-21.
CrossRefDirect Link - Jaccard, P., 1908. Nouvelles researches sur la distribution florale. Bull. Soc. Vaudoise Sci. Natl., 44: 223-270.
CrossRefDirect Link - Ogunbayo, S.A., D.K. Ojo, R.G. Guei, O.O. Oyelakin and K.A. Sanni, 2005. Phylogenetic diversity and relationship among forty rice accessions using Morphological and RAPDs techniques. Afr. J. Biotechnol., 4: 1234-1244.
Direct Link - Sasaki, T., 2002. Rice genomics to understand rice plant as an assembly of genetic codes. Curr. Sci., 83: 834-839.
Direct Link - Virk, P.S., J. Zhu, G.J. Newbury, M.T. Jackson and B.V. Ford-Lloyd, 2000. Effectiveness of different classes of molecular marker for classifying and revealing variation in rice (Oryza sativa) germplasm. Euphytica, 112: 275-284.
Direct Link