
Shoukat Hussain Shah
Department of Botany, Aligarh Muslim University, Aligarh-202 002, India
Asian Journal of Plant Sciences
Year: 2007 | Volume: 6 | Issue: 2 | Page No.: 403-406







ABSTRACT
The effects of various levels of basal nitrogen (0, 176, 264, 352 or 442 mg N pot-1) applied with or without 10-5 M kinetin (KIN) spray were analysed through a pot experiment conducted on Nigella sativa L. Although, N alone was found to significantly enhance all parameters, (viz., nutrient (NPK) accumulation, number of capsules, seed yield plant-1, oil and essential oil yields plant-1 except oil and essential oil contents. An additional treatment of KIN further improved all the elevated values synergistically. Moreover, the effect of KIN was most pronounced along with 325 mg N pot-1, as compared to other combined treatments.
PDF Abstract XML References Citation
How to cite this article
Shoukat Hussain Shah, 2007. Influence of Combined Application of Nitrogen and Kinetin on Nutrient Uptake and Productivity of Black Cumin (Nigella sativa L.). Asian Journal of Plant Sciences, 6: 403-406.
DOI: 10.3923/ajps.2007.403.406
URL: https://scialert.net/abstract/?doi=ajps.2007.403.406
DOI: 10.3923/ajps.2007.403.406
URL: https://scialert.net/abstract/?doi=ajps.2007.403.406
INTRODUCTION
Black cumin (Nigella sativa L.) is a plant of repute in the Greco-Arab Eastern and Islamic systems of traditional medicine, where it finds immense use as a carminative, emanagogue, galoactagogue, digestive, stimulant and diaphoretic (Riaz et al., 1996). The ripe seed and its oil effectively alleviates pityriasis, leucoderma, ringworm, eczema, chest congestion, migraine, paralysis and rheumatism. Despite of its immense medical and aromatic importance, this herb has escaped cultivation and has naturalized in and around the Middle East. Its pressing demand is hence, met primarily through collection from forests and even the most focused attempts at its large scale cultivation have met with little success. This is because, there is a dearth of information regarding nutritional techniques that can be adopted to pioneer and enhance its productivity.
Nitrogen is the single most important limiting nutrient for crop growth (Goyal and Huffaker, 1984) and hence ample N availability acts as a key contributing factor towards wholesome improvement in agricultural production (Shah, 2004). However, taking into consideration the ill effects of employing large amounts of inorganic fertilizers, the use of supplementary treatments such as phytohormone sprays can be said to hold considerable practical relevance. The application of cytokinins has been shown to appreciably enhance crop productivity and performance (Mousa et al., 2001; Shah, 2006; Shah et al., 2006). Besides exogenous KIN spray has also been found to increase activities of nitrate reductase (Roa et al., 1984; Saxena and Saxena, 2002), glutamate synthetase (Ghisi and Passera, 1987) and enhance chlorophyll and protein contents (Towne and Owensby, 1983). CKs may also be employed to improve the photosynthetic rate and the activity of RuBPCO (Singh et al., 2001).
In this context, it was therefore thought relevant to study the responses of Nigella sativa L. to application of various N fertilization dosages, with or without spray of 10-5 M KIN.
MATERIALS AND METHODS
A pot experiment was carried out on Nigella sativa L. during the winter season of 2001-2002, at the Department of Botany, Aligarh Muslim University, Aligarh, situated at 27°52´ N latitude, 78°51´ E longitude and 187.5 m above the sea level. Seeds were obtained from the Regional Research Institute of Unani Medicine, Aligarh, India and were surface sterilized with 0.01% mercuric chloride solution, followed by repeated washings using double distilled water. They were then sown in the earthen pots (25 cm diam) filled with soil and farmyard manure, mixed in a ratio of 9:1. Each treatment was replicated thrice. The soil of the experimental pots was sandy loam in texture and available N was 185 Kg N ha-1. The basal application of N in the form of urea at the rate (0, 176, 264, 352 or 442 mg N pot-1) and the administration of 10-5 M KIN spray at the rate of 5 cm3 plant-1 was done at 40 DAS (vegetative stage). The concentration of KIN is based on a previous experiment (Shah et al., 2006).
After 70 days of sowing (DAS), the plant samples were oven dried, ground and chemically analysed for N and P contents, adopting the methods of Lindner (1944) and Fiske and Subba Row (1925), respectively. Content of K was analysed by flame photometer. The uptake of N, P and K were the product of their concentration in the plant and dry weight of plant-1. Five plants from each treatment were selected and capsule number plant-1 was recorded. The seed yield from five randomly selected plants was noted after threshing the seeds. The seed oil content was determined according to the method recommended by AOAC (1970). Extraction of essential oil was done with 30 g seed sample following hydrodistillation for 2 h. Analysis of variance was carried out on the data obtained and LSD (p = 0.05) was calculated as per Gomez and Gomez (1984).
RESULTS AND DISCUSSION
Nutrient (NPK) uptake of N. sativa L. was found to be favourably influenced by the application of inorganic N and increased with increasing N dosage, being maximum at 352 mg N pot-1. A higher level i.e., 442 mg N pot-1 proved supra-optimal and was less effective (Table 1). N is a key constituent of proteins, which inturn provide a basic foundation for the plant infrastructure (Shah, 2004). Implicably, ample availability of N to a plant during its growth phase results in efficient utilization and manifestation of this basic nutrient in the plant stature. Hence, it can be proposed that the cause for the increase observed in nutrient uptake may be the increased assimilation of all nutrients under maximum uptake potential, generated by maximal growth (data not shown). These results are ascertained by those of Gastal and Lemarie (2002) and Lawlor (2002). Synchronously, KIN was found to further contribute to an increase in the nutrient uptake, which might have been due to the phytohormone induced growth and development of roots, hence allowing further exploration of the soil substrates (Simpson et al., 1982). Another probable cause may be the increase in transpiration due to KIN (Pospíšilová et al., 2000) thereby, encouraging mass flow to the roots and facilitating nutrient uptake. Similar effects of KIN have also been observed by Kannan and Mathew (1970), Simpson et al. (1982) and Rangacharya and Bawankar (1991).
Number of capsules and seed yield plant-1 were found to be appreciably enhanced by the fertilizer treatment, with result being most pronounced for application of 352 mg N pot-1 (Table 2). Here again, treatment with KIN along with the optimum dosage of N was found to considerably boost the overall yield (Table 2). Proposedly, the increase in NPK accumulation in the test plants must have highly optimized the nutrient availability, ultimately resulting on enhancement of vegetative and reproductive growth (data not shown). This might have caused an increase in photosynthate production, as well as its translocation from the source to sink (capsules/pods), thereby increasing their number (Dybing, 1994) and subsequently elevating seed yield (Table 2). Such increase corroborates the findings of Uppar and Kulkarni (1989) and Azam (2002). Likewise, the observed increase in aforesaid parameters due to KIN may be ascribed to the increased opportunity for formation and development of new buds (Bruinsma, 1977), coupled with successful prevention of senescence (Nagel et al., 2001), thereby causing more available yield at maturity.
In the present study oil content of seeds was found to decrease with increasing levels of N, as well as, with KIN (Table 2). The probable reason for this adverse effect of the treatment may be the preferential utilization of C-skeletons at the time of seed filling towards protein synthesis (data not shown), rather than oil formation (Shah, 2004). However, owing to the increased seed yield, the oil yield obtained after the combined treatment, especially with 352 mg N pot-1, was so spectacular (Table 2) that it outbalanced the lowered oil content-an obvious commercial advantage of the seeds.
| Table 1: | Effect of various levels of basal N (mg pot-1) in combination with the foliar spray of 10-5 M KIN on NPK uptake of Nigella sativa L. at 70 DAS |
 | |
| aW0, Control | |
| Table 2: | Effect of various levels of basal N (mg pot-1) in combination with the foliar spray of 10-5 M KIN on capsule number, seed yield, oil content, oil yield, essential oil content and essential oil yield plant-1 of Nigella sativa L. at maturity (130 DAS) |
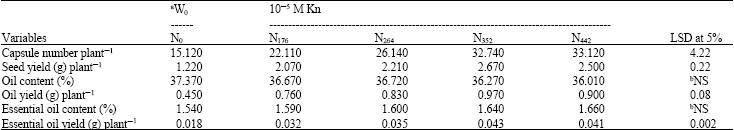 | |
| aW0, Control, bNS: Not Significant | |
These results are on expected lines and corroborate those of Omer et al. (1995), Singh (2001) and Hashmi (2003).
Further, results indicated that application of N or phytohormone showed non-significant increase in the essential oil content over the respective control (Table 2). This may well be attributed to the dilution with growth effect, as the sink became more voluminous due to treatment with optimum level of N (352 mg N pot-1) and KIN. This fact is corroborated by the high essential oil yield plant-1 (Table 2), thus confirming the work of Mousa et al. (2001) and Hashmi (2003).
CONCLUSION
It is therefore concluded that the application of basal N at the rate of 352 mg N pot-1, under administration of foliar 10-5 M KIN enabled maximum exploitation of soil resources, hence resulting in enhanced overall yield of N. sativa L.
ACKNOWLEDGMENT
The author wish to thank Dr. A. Ridal for his critical reading of the manuscript and valuable comments.
REFERENCES
- Dybing, C.D., 1994. Soybean flower production as related to plant growth and seed yield. Crop Sci., 34: 489-497.
Direct Link - Fiske, C.H. and Y. Subbarow, 1925. The colorimetric determination of phosphorus. J. Biol. Chem., 66: 375-400.
CrossRefDirect Link - Gastal, F. and G. Lemarie, 2002. N uptake and distribution in crops and agronomical and eco-physiological perspective. J. Exp. Bot., 53: 789-799.
CrossRef - Gomez, K.A. and A.A. Gomez, 1984. Statistical Procedures for Agricultural Research. 2nd Edn., John Wiley and Sons Inc., Hoboken, New Jersey, ISBN: 978-0-471-87092-0, Pages: 704.
Direct Link - Kannan, S. and T. Mathew, 1970. Effects of growth substances on the absorption and transport of iron in plants. Plant Physiol., 45: 206-209.
PubMed - Lawlor, D.W., 2002. Carbon and nitrogen assimilation in relation to yield: Mechanisms are the key to understanding production systems. J. Exp. Bot., 53: 773-787.
CrossRefDirect Link - Mousa, G.T., I.H. El-Sallami and E.F. Ali, 2003. Response of Nigella sativa L. to foliar application of gibberellic acid, benzyladenine, iron and zinc. Assiut. J. Agric. Sci., 32: 141-156.
Direct Link - Nagel, L., R. Brewster, W.E. Riedell and R.N. Reese, 2001. Cytokinin regulation of flower and pod set in soybeans (Glycine max (L.) Merr.). Ann. Bot., 88: 27-31.
Direct Link - Pospisilova, J.H. Synkova and J. Rulcova, 2000. Cytokinins and water stress. Biol. Plant., 43: 321-328.
Direct Link - Riaz, M., M. Syed and F.M. Chaudhary, 1996. Chemistry of the medicinal plants of the genus Nigella (family-Ranunculaceae). Hamdard Medicus, 39: 40-45.
Direct Link - Roa, L.V.M., N. Datta, M. Mahadevan, S. Guha-Mukherjee and S.S. Sopory, 1984. Influence of cytokinins and phytohrome on nitrate reductase activity in etiolated leaves of maize. Photochemistry, 23: 1875-1879.
CrossRef - Saxena, A. and D.K. Saxena, 2002. Nitrate reductase activity and chlorophyll content in Sphagnum as affected by kinetin, glutathione and metals. Indian J. Plant Physiol., 7: 82-85.
Direct Link - Shah, S.H., I. Ahmad and Samiullah, 2006. Responses of Nigella sativa L. to foliar application of gibberellic acid and kinetin. Biol. Plant., 51: 563-566.
Direct Link - Simpson, R.J., H. Lambers and J. Michael, 1982. Kinetin application to roots and its effect on uptake, translocation and distribution of nitrogen in wheat (Triticum aestivum) grown with a split root system. Physiol. Plant, 56: 430-435.
CrossRef - Singh, D.V., G.C. Srivastava and M.Z. Abdin, 2006. Amelioration of negative effect of water stress in Cassia angustifolia by benzyladenine and/or ascorbic acid. Biologia Plantarum, 44: 141-143.
CrossRefDirect Link - Towne, G. and C. Owensby, 1983. Cytokinins effect on protein and chlorophyll content of big bluestem leaves. J. Range Manage., 36: 75-77.
Direct Link - Shah, S., 2007. Effect of kinetin spray on growth and productivity of black cumin plants. Russian J. Plant Physiol., 54: 702-705.
CrossRefDirect Link