
George N. Chemining`wa
Not Available
J.W. Muthomi
Not Available
S.W.M. Theuri
Not Available
Asian Journal of Plant Sciences
Year: 2007 | Volume: 6 | Issue: 7 | Page No.: 1113-1118







ABSTRACT
A study was conducted in Kenya at the University of Nairobi’s Field Station in 2004 long and short rains to determine the effect of rhizobia inoculation and starter-N on nodulation, shoot biomass and yield of selected grain legumes. Common bean (Phaseolus vulgaris L.), lima bean (Phaseolus lunatus L.), cowpea (Vigna unguiculata L.), green gram (Vigna radiata L.), pigeon pea (Cajanus cajan L.) and lablab (Lablab purpureus L.) were uninoculated, inoculated with appropriate rhizobia, or supplied with 26 kg N ha-1 as starter-N. Rhizobia inoculation had no effect on nodule numbers in both seasons. Starter-N suppressed nodule numbers only in the long rains at 4 weeks after emergence. In general, common bean had higher nodule numbers and nodule biomass than other legumes while lima bean registered fewest nodules. Rhizobia inoculation improved nodule biomass in short rains at 6 weeks after emergence, while starter-N had no effect on nodule biomass in both seasons. Inoculation and starter-N had no effect on shoot biomass and grain yield in both seasons. Lablab, common bean and lima bean had higher shoot biomass than other legumes in both seasons. In long rains, lima bean produced highest grain yield followed by common bean while the converse was the case in short rains. Pigeon pea and green gram performed poorest in short rains. It was concluded that it might not be necessary to inoculate or apply starter-N to legumes under soil conditions prevailing in the current study.
PDF Abstract XML References Citation
How to cite this article
George N. Chemining`wa, J.W. Muthomi and S.W.M. Theuri, 2007. Effect of Rhizobia Inoculation and Starter-N on Nodulation, Shoot Biomass and Yield of Grain Legumes. Asian Journal of Plant Sciences, 6: 1113-1118.
DOI: 10.3923/ajps.2007.1113.1118
URL: https://scialert.net/abstract/?doi=ajps.2007.1113.1118
DOI: 10.3923/ajps.2007.1113.1118
URL: https://scialert.net/abstract/?doi=ajps.2007.1113.1118
INTRODUCTION
Crop production in most smallholder farms in Kenya is characterized by continuous cultivation coupled with low input use, which results in reduced native soil fertility and productivity (Wahome, 2007; Ojiem et al., 2000; Lynam et al., 1998). Nitrogen is one of the most limiting nutrients in these farms for crop production. Inorganic N fertilization is needed to alleviate N deficiency. Nitrogen fertilization, however, is costly and therefore out of reach of most resource-poor farmers. Manure obtained from livestock could be used as a cheap source of nutrients, but nutrient contents are often lower, which requires bulk application to satisfy plant nutrient demand (Kihanda, 1996). Inclusion of legumes into existing cropping systems has the potential to reduce or eliminate inorganic N fertilization due to their ability to fix N from the atmosphere (Giller, 2001; Houngnandan et al., 2000). Reports indicate that legumes can fix as much as 200 kg N ha-1 year-1 under optimal field conditions (Giller, 2001; Rowland, 1993). Such fixation of N can only be achieved in the presence of efficient rhizobial strains, which can be native to the soil or introduced in-form of commercial inoculants. Inoculation of legumes with effective rhizobia can improve grain yields (Nkwiine and Rwakaikara-Silver, 2007; Zengenia et al., 2006; Giller, 2001). Inoculation is, however, not universal and inoculation does not always elicit positive responses. Inoculation of legumes is necessary in absence of compatible rhizobia and when rhizobial populations are low or inefficient in fixing N (Abaidoo et al., 2007; Fening and Danso, 2002; Catroux et al., 2001; Brockwell et al., 1995). Awareness and use of rhizobia inoculants in legume production in Kenya is limited (Wahome, 2007; Woomer et al., 1997). Most of the rhizobia inoculation studies conducted in Kenya have concentrated on common bean. Little attention has been paid on other grain legumes such as green gram, lima bean, cowpea and lablab which have the potential to broaden the food base and thereby improve food security. In such legumes, the effect of inoculation on crop yields has not been adequately addressed. The objective of this study was to investigate the effect of rhizobia inoculation and starter-N application on nodulation and yields of grain legumes.
MATERIALS AND METHODS
Experimental site: The study was conducted at University of Nairobi’s Faculty of Agriculture Field Station in Nairobi, Kenya. The site is located at latitude 01° 15’S and longitude 36° 44’ E, 1,850 m above sea level. The site experiences bimodal rainfall pattern, with long rains received between March and May and the short rains received between October and December every year. The average annual rainfall is about 1000 mm. Soils of the area are deep, well-drained, dark reddish brown to dark red friable clay with acidic humic topsoil (humic NITOSOLS) of moderate fertility developed from Limuru Trachite (Michieka, 1977). Daily temperature ranges from 12 to 28°C with a mean of about 20°C.
Initial soil N, P and soil organic carbon levels at the experimental site were about 2 g kg-1, 20 mg kg-1 and 35 g kg-1, respectively. The population size of indigenous cowpea rhizobia (nodulating cowpea, pigeon pea, green gram, lima bean and lablab) and common bean rhizobia in the experimental site averaged 3.1x102 and >9.0x102 bacterial cells g-1 soil, respectively (Chemining’wa and Vessey, 2006). The weather conditions at the study site are shown in Fig. 1 and 2.
Experimental design, treatments and crop husbandry: The field experiments were conducted for two consecutive seasons: long rains from March to August 2004 and short rains from October to January 2005 (Fig. 1, 2). The first season and second season crops were grown in different plots on the same field. The test legume crops were lima bean (local variety), pigeon pea (variety Mbaazi 1), green gram (variety N26), cowpea (variety M66), lablab variety (DH1002) and common bean (variety GLP-2). The treatments consisted of rhizobia inoculation, starter-N fertilizer application at 26 kg N ha-1 and a control (0 kg N ha-1 and no rhizobia inoculation). A Randomized Complete Block Design (RCBD) with a split plot arrangement of treatments was used in both seasons and each treatment was replicated three times. Legume species were assigned to the main plots and the rhizobia inoculation, starter-N fertilization and control treatments were assigned to the subplots. Phosphorous was applied to all plots at planting at 20 kg P ha-1. Starter N was applied in form of calcium ammonium nitrate. Fertilizers were banded along crop rows to a depth of 15 cm to avoid direct contact with seeds. Rhizobia inocula were obtained from the MIRCEN Project, University of Nairobi.
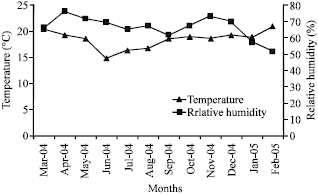 | |
| Fig. 1: | Rainfall distribution at University of Nairobi’s Field Station study site |
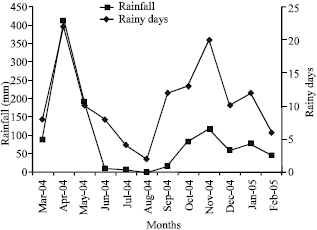 | |
| Fig. 2: | Temperature and relative humidity at University of Nairobi’s Field Station study site |
Rhizobium leguminosarum bv. viciae strain 2674 was used to inoculate common bean while cowpea cross-nodulating Bradyrhizobium sp. strain 3456 was used to inoculate pigeon pea, cowpea, lima bean, green gram and lablab. Seeds were inoculated with peat-based inoculants containing gum Arabic adhesives to ensure good contact with the inoculant. Care was taken to avoid cross contamination of inoculated and uninoculated seeds by planting the uninoculated seeds prior to inoculated seeds. The size of each experimental plot was 3.0x2.0 m with an inter-plot spacing of 1 m. Legume seeds for the following species were planted at the recommended spacing: common bean 30x15 cm, pigeon pea 50x20 cm, lima bean 40x25 cm, green gram 30x10 cm, lablab 80x50 cm and cowpea 75x20 cm. The experimental fields were kept relatively weed-free by hand weeding.
Data collection and analysis: Sampling for number of nodules, nodule dry matter and shoot biomass for all the treatments was conducted at 4 and 6 WAE in both seasons. Three plants were carefully uprooted from each experimental plot by digging 15 cm around the plant using a spade and washed with clean tap water to remove all attached soil from the roots and the nodules. The nodules were counted and carefully picked using a pair of tweezers, oven dried at 60°C for 48 h and weighed. The shoots of the plants were oven-dried at 60°C for 72 h and dry matter weights determined. At physiological maturity, pods were harvested from each experimental plot, excluding the outer rows and the outer guard plants in each row and shelled. The grains were sun-dried and weighed. Analysis of variance (ANOVA) was performed on the data using Genstat 5 Release 3.2 package (Genstat, 1995) and means separated by the Least Significant Difference (LSD) test at 5% probability level (Steel and Torrie, 1987).
RESULTS AND DISCUSSION
Rhizobia inoculation did not significantly (p≤0.05) improve nodule number plant-1 at 4 and 6 weeks after emergence (WAE) in both seasons (Table 1, 2). Starter N suppressed nodulation by 41% during the long rains at 4 WAE. In both seasons, common bean produced significantly more nodules per plant than most of the other legumes while lima bean generally registered fewer nodules than the rest of the legumes.
Rhizobia inoculation improved nodule biomass by 33% in the short rains but had no significant (p≤0.05) effect on this parameter in the long rains (Table 3). In the long rains, lablab and common bean accumulated significantly the highest amount of nodule dry matter compared to the rest of the legumes. In the short rains, lablab out-performed common bean in nodule biomass while lima bean and pigeon pea had the lowest nodule biomass.
Rhizobia inoculation and starter-N treatments did not significantly (p≤0.05) improve shoot biomass plant-1 and grain yield in both seasons (Table 4, 5). In general, lablab, common bean and lima bean had significantly (p≤0.05) higher shoot biomass than the other legumes (Table 4). In the long rains, lima bean out-yielded common bean by 70% which in turn out-yielded cowpea, lablab, green gram and pigeon pea by 164, 120, 341 and 484%, respectively (Table 5). In the short rains, common bean out-yielded lima bean by 25%, which in turn out-yielded cowpea by 23%. Cowpea out-yielded lablab by 37% which also out-yielded green gram and pigeon pea by 585 and 879%, respectively (Table 5).
Rhizobia inoculation did not improve the nodule numbers in both seasons, suggesting that the rhizobial population densities nodulating the test legumes were adequate in the trial fields. Thies et al. (1991) showed that inoculation of eight leguminous crops growing in soils containing 10 to 100 indigenous rhizobial cells g-1 soil increased the number of nodules per plant. In the present study, the population size of indigenous rhizobia nodulating cowpea was 3.1x102 rhizobia cells g-1 dry soil while that of common bean was above 9.0x102 rhizobia cells g-1 dry soil (Chemining’wa and Vessey, 2006). These high rhizobia population levels most likely prevented nodulation response to inoculation among the legumes. Establishment of inoculant strains in soils with substantial populations of indigenous rhizobia is considered difficult and response to inoculation unlikely (Abaidoo et al., 2007; Bloem and Law, 2001; Houngnandan et al., 2000). Starter-N caused a decline in nodule numbers in the short rains. High levels of inorganic N, especially nitrate-N, have been shown to suppress nodulation of legumes (Chemining’wa and Vessey, 2006). However, under soils low in mineral N, a moderate dose of starter-N has been demonstrated to stimulate seedling growth and subsequently N2-fixation (Goi et al., 1993). Inorganic N is required by legume plants during the ‘nitrogen hunger period’ for their nodule development, shoot and root growth before the onset of N2-fixation process (Hansen, 1994). Nodulation was not improved by application of 26 kg N ha-1 possibly because the initial soil N level (20% N) was adequate during the nitrogen hunger period. Nodule biomass responded positively to rhizobial inoculation during the short rains at 6 WAE, suggesting that the inoculant strain was possibly more efficient than indigenous strains. Nodulation varied with legume species. Among the legumes, common bean plants were the most nodulated while lima bean plants were the least nodulated. This could be attributed to genetic differences among the species as interactions between legume species and microsymbionts have been demonstrated to be highly specific (Qiang et al., 2003). This suggests that bradyrhizobia strains in the study site did not effectively nodulate lima bean that belongs to the same cross-inoculation group with cowpea, green gram, pigeon pea and lablab. In trials set up in 10 geographically diverse sites in Kenya, Mureithi et al. (1998) found higher nodule numbers per plant for cowpea (27.3), pigeon pea (16) and lablab (6) than for lima bean (1.7).
| Table 1: | Nodule number plant-1 of grain legumes at 4 weeks after emergence during the long and short rains at Kabete in 2004 |
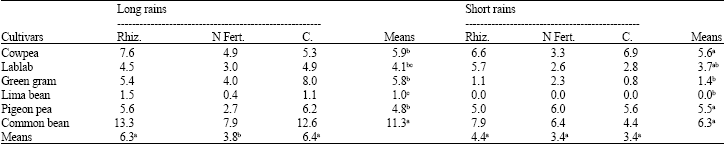 | |
| Rhiz. = Rhizobia; N Fert. = N Fertilizer; C = Control, Means followed by the same letter(s) within a row or a column are not significantly different at the p = 0.05 level using LSD test | |
| Table 2: | Nodule number plant-1 of grain legumes at 6 weeks after emergence during the long and short rains at Kabete in 2004 |
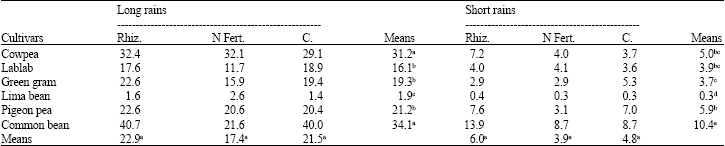 | |
| Rhiz. = Rhizobia; N Fert. = N Fertilizer; C = Control, Means followed by the same letter(s) within a row or a column are not significantly different at the p = 0.05 level using LSD test | |
| Table 3: | Nodule dry matter (mg plant-1) of grain legumes at 6 weeks after emergence during the long and short rains at Kabete in 2004 |
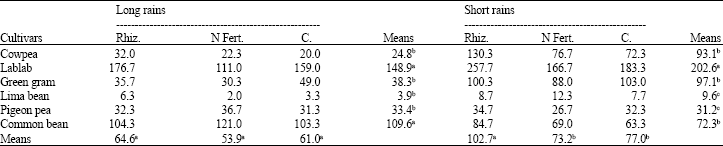 | |
| Rhiz. = Rhizobia; N Fert. = N Fertilizer; C = Control, Means followed by the same letter(s) within a row or a column are not significantly different at the p = 0.05 level using LSD test | |
| Table 4: | Shoot dry matter (g plant-1) of grain legumes at 6 weeks after emergence during the long and short rains at Kabete in 2004 |
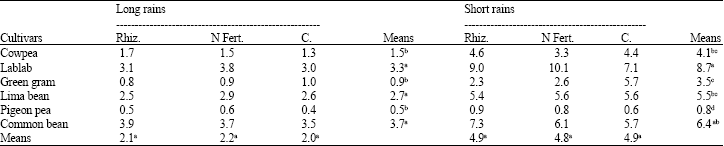 | |
| Rhiz. = Rhizobia; N Fert. = N Fertilizer; C = Control, Means followed by the same letter(s) within a row or a column are not significantly different at the p = 0.05 level using LSD test | |
| Table 5: | Grain yield (kg ha-1) of legume species during the long and short rains at Kabete in 2004 |
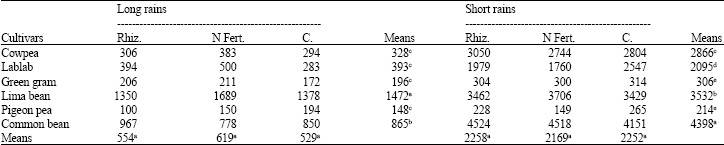 | |
| Rhiz. = Rhizobia; N Fert. = N Fertilizer; C = Control, Means followed by the same letter(s) within a row or a column are not significantly different at the p = 0.05 level using LSD test | |
Rhizobia inoculation failed to improve shoot biomass and yield in all the grain legumes. Lack of response to Rhizobium inoculation has also been reported in common bean studies conducted in low-N sites in western Kenya (Musandu and Ogendo, 2002). This was attributed to the abundance of ineffective indigenous rhizobia. The grain yield achieved in the present study was dependent on the legume species. Lima bean and common bean had the highest grain yield over the two seasons. Lima bean performed particularly well during the long rain season that received poorly distributed rains. This could be attributed to its adaptation to water stress (Rowland, 1993). Green gram and pigeon pea produced the lowest grain yield compared to the other legumes over the two seasons, suggesting that varieties used in the study may not be suitable for the low temperature conditions prevailing at the experimental site.
The lack of nodulation response to inoculation in grain legumes could be attributed to poor inoculant viability, adequate soil mineral N, incompatibility of the inoculant strain with specific grain legumes or the presence of highly competitive native rhizobia that restricted occupancy of the nodules by the inoculant strains. Inoculation and starter-N did not improve yields, suggesting that it may not be necessary for farmers with similar soil conditions to inoculate or apply starter-N with the aim of improving yield. It would, however, be worthwhile to conduct a similar study in N-depleted fields prevalent in smallholder production systems.
ACKNOWLEDGMENT
The African Institute for Capacity Development (AICAD) is gratefully acknowledged for funding this research.
REFERENCES
- Abaidoo, R.C., H.H. Keyser, P.W. Singleton, K.E. Dashiell and N. Sanginga, 2007. Population size, distribution and symbiotic characteristics of indigenous Bradyrhizobium sp. that nodulate TGx soybean genotypes in Africa. Applied Soil Ecol., 35: 57-67.
CrossRef - Bloem, J.F. and I.J. Law, 2001. Determination of competitive abilities of Bradyrhizobium japonicum strains in soils from soybean production regions in South Africa. Biol. Fert. Soils, 33: 181-189.
Direct Link - Brockwell, J., P.J. Bottomley and J.E. Thies, 1995. Manipulation of rhizobia microflora for improving legume productivity and soil fertility: A critical assessment. Plant Soil, 174: 143-180.
Direct Link - Catroux, G., A. Hartmann and C. Revelin, 2001. Trends in rhizobial inoculant production and use. Plant Soil, 230: 21-30.
CrossRef - Chemining`wa, G.N., M.D.G. Njarui, J.G. Mureithi, J.N. Gitari and S.N. Maobe, 2006. Response of Green Manure Legumes to Phosphorous Application and Rhizobial Inoculation. In: Enhancing Agricultural Productivity in East Africa; Development and Up-scaling of Green Manure Legume Technologies in Kenya, Mureithi, J.G., C.K.K. Gachene, J.W. Wamuongo and M. Eilitta (Eds.). Kenya Agriculture Reserch Institute, Nairobi, pp: 45-60.
- Chemining`wa, G.N. and J.K. Vessey, 2006. The abundance and efficacy of Rhizobium leguminosarum bv. viciae in cultivated soils of the eastern Canadian prairie. Soil Biol. Biochem., 38: 294-302.
Direct Link - Fening, J.O. and S.K.A. Danso, 2002. Variation in symbiotic effectiveness of cowpea bradyrhizobia indigenous to Ghanaian soils. Applied Soil Ecol., 21: 23-29.
CrossRef - Goi, S.R., J.I. Sprent, E.K. James and J. Jacob-Neto, 1993. Influence of nitrogen form and concentration on the nitrogen fixation of Acacia auriculiformis. Symbiosis, 14: 115-122.
Direct Link - Tang, C., P. Hinsinger, B. Jaillard, Z. Rengel and J. Drevon, 2001. Effect of phosphorus deficiency on the growth, symbiotic N2 fixation and proton release by two bean (Phaseolus vulgaris) genotypes. Agronomy, 21: 683-689.
CrossRefDirect Link - Houngnandan, P., N. Sanginga, P. Woomer, B. Vanlauwe and O. van Cleemput, 2000. Response of Mucuna pruriens to symbiotic nitrogen fixation by rhizobia following inoculation in farmers fields in the derived savanna of Benin. Biol. Fert. Soils, 30: 558-565.
Direct Link - Musandu, A.A.O. and J.O. Ogendo, 2002. Effect of Rhizobium Inoculation and Fertilizer Nitrogen and Phosphorous on the Grain Yield of Beans in Western Kenya. Challenges and Imperatives for Biological Nitrogen Fixation Research and Application in Africa for the 21st Century, Karanja, N.K. and H.P. Kahindi (Eds.). John Philips Africa Limited, Nairobi,.
CrossRef - Nkwiine, C. and M.C. Rwakaira-Silver, 2007. Status of research on soil microsymbionts in Uganda. Afr. J. Ecol., 45: 27-35.
Direct Link - Ojiem, J.O., J.G. Mureithi and E.A. Okuosa, 2000. Integrated Management of Legume Green Manure, Farmyard Manure and Inorganic Nitrogen for Soil Fertility Improvement in Western Kenya. In: Participatory Technology Development for Management by Smallholders in Kenya, Mureithi, J.G., C.K.K. Gachene, F.N. Muyekho, M. Onyango, L. Mose and O. Magenya (Eds.). Agricultural Research Institute, Nairobi, Kenya, pp: 97-102.
- Qiang, C., Z. Xiapong, T. Zewdi, K. Seppo, L. Dengyu, and L. Kristina, 2003. Diversity of peanut (Arachis hypogaea) bradyrhizobia and their host plants. Plant Soil., 255: 605-617.
Direct Link - Thies, J.E., P.W. Singleton and B.B. Bohlool, 1991. Modeling symbiotic performance of introduced rhizobia in the field by use of indices of indigenous population size and nitrogen status of the soil. Applied Environ. Microbiol., 57: 29-37.
Direct Link - Zengeni, R., S. Mpepereki and K.E. Giller, 2006. Manure and soil properties affect survival and persistence of soybean nodulating rhizobia in smallholder soils of Zimbabwe. Applied Soil Ecol., 32: 232-242.
Direct Link