
M. Tariq
Not Available
C.J.B. Mott
Not Available
Asian Journal of Plant Sciences
Year: 2006 | Volume: 5 | Issue: 6 | Page No.: 963-969







ABSTRACT
The present study was based on the hypothesis that B has a retarding influence on the accumulation of anions in plants. A study was carried out to assess the influence of B on the concentration and uptake of anions in radish (cv. French breakfast) crop, using sand culture technique, under green house conditions. The experiment was laid out in a randomized complete block design and replicated three times. Boron was applied at the rate of 0, 0.25, 0.50, 1.0, 2.0, 3.0 and 5.0 mg B L-1 as H3BO3 along with a basal dose of modified complete nutrient solution based on the Long Ashton Formula. Results revealed that significant treatment effects were found on the growth response of radish plants and maximum yield was recorded at 0.5 mg L-1 of added B. Toxic effects accompanied by considerable yield decreases was observed at higher levels of B supply. The concentration of P and B in plants were increased, S decreased and N remain unchanged. While, the total uptake of all anions except B significantly decreased with increasing levels of B in the nutrient solution and showed close similarity to the growth response of radish plants, suggesting that total uptake of anions was useful but alternative way of estimating the nutrient behavior with regard to B supply. Generally, low and high levels of added B had detrimental effects on the concentration and total uptake of anions in radish plants. The present study suggests that B supply had specific effects with respect to different anions.
PDF Abstract XML References Citation
How to cite this article
M. Tariq and C.J.B. Mott, 2006. Influence of Boron on the Concentration and Uptake of Anions by Radish (Raphanus sativus L.). Asian Journal of Plant Sciences, 5: 963-969.
DOI: 10.3923/ajps.2006.963.969
URL: https://scialert.net/abstract/?doi=ajps.2006.963.969
DOI: 10.3923/ajps.2006.963.969
URL: https://scialert.net/abstract/?doi=ajps.2006.963.969
INTRODUCTION
Boron is of course, one of the recognized micronutrients for plant growth and production. A great deal of attention has been given to studying the effects of B supply on the accumulation of other nutrient-elements, specifically cations (K, Ca, Mg and Na), but information on the concentration and uptake of anions (N, P and S) is limited. Probably, Rehim (1937) was the first pioneer, who found that the addition of B to the nutrient medium increased the intake of cations and retarded the anions in plants as compared to culture lacking B. Similarly, Batey (1971) reported that B is thought to have a favorable influence on the absorption of cations particularly Ca, to have retarding influence on the absorption of anions and to have an essential part in carbohydrate and N metabolism. In the same year this was also confirmed by Valmis and Ulrich (1971) who reported that with increasing B in the nutrient solution the concentrations of N, P and S linearly decreased in the leaves of sugar beet. Bonilla et al. (1980) observed that both B deficiency and toxicity resulted in more NO3-N accumulation in the sap of sugar beet due to the decrease in the activity of the N-Rase enzyme, suggesting a specific effect of B on N-Rase activity. Pollard et al. (1977) found that B deficiency in corn and broad beans reduced the capacity for the absorption of PO4, due to the reduced ATPase activity, which could be rapidly restored by the addition of B. Gupta and Sanderson (1993) reported non- significant interactions between S and B in potato crop. It is evident from the literature that the deficiency or excess of B may affect the anion values in plants. However, information regarding the effect of B on the availability of anions is limited in the literature and the nature of these complex interactions between B and other anions, such as N, P and S are still obscure. Therefore, the present study was undertaken to test the general hypothesis, that B has a retarding influence on the accumulation of anions in plants, using sand culture technique with the main objective, to assess the influence of B on the concentration and uptake of anions by radish plants.
MATERIALS AND METHODS
Experimental conditions and design: The experiment was carried out in the green house. The day/night temperature varied with in the range 22 to 18°C, respectively. Artificial illumination was used to give 16 h day-1 and the relative humidity was around 65% during the experiment. The experiment was laid out in a randomized complete block design and replicated three times. The three blocks together therefore, gave a total of 21 pots. Each block was situated within a distance of 30 cm of each other. The position of each pot was randomly changed once a week, to minimize the spatial variations in the green house during the course of experiment.
Sowing: Six radish seeds (cv. French breakfast) was sown uniformly 1 cm deep and 2.5 cm apart from one another, in each 15 cm plastic pot containing 1 kg of acid leached fine sand and the surface of sand covered with black alkathene granules, to prevent rapid loss of moisture and algal growth. A glass wool filter paper was placed at the bottom of each pot to cover holes and the pot placed on a plastic saucer. The moisture content of the sand was kept at approximately 60% of its water holding capacity. The seeds germinated within a week and upon establishment the seedlings were thinned out so that finally each pot contained four equal size radish plants.
Basal nutrient and boron solutions: All the nutrient solutions were prepared from AnalaR Grade chemicals and deionised water of a conductivity 0.20 μmhos cm-1. Boron was applied at the rate of 0, 0.25, 0.50, 1.0, 2.0, 3.0 and 5.0 mg B L-1 as H3BO3 along with a basal dose of modified complete nutrient solution based on the Long Ashton Formula as recommended by Hewitt (1966). The detail of the various salts concentration used as shown in Table 1. Complete nutrient solution with different B concentrations were started a week after germination. The nutrient solutions were freshly prepared whenever added and the pH of each solution was maintained to 5.5±0.1 either with 0.1 M HCl or 0.1 M NaOH. Each pot was kept at a constant moisture content by means of alternate day additions of culture solution following weighing of the pot. A total quantity of 200 mL was supplied to each pot during the course of the experiment. During the final week of the experiment the plants only received deionised water.
| Table 1: | Chemical composition and concentration of basal nutrient solution, based on (modified) Long Ashton Formula |
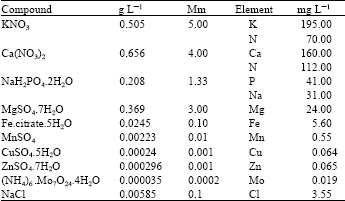 | |
Stock solutions for each nutrient element were prepared separately in the plastic volumetric flasks and stored in a refrigerator. At each B level the nutrient solution was prepared by mixing the appropriate volume of concentrated nutrient stock solutions.
Water loss evaluation: Water losses by evapotranspiration was monitored by weighing daily the control pots (with out seedlings) as well as the varying B treatment pots. It is evident that the moisture content of the sand depended on the intensity of evapotranspiration of pots i.e. growth rate, temperature and relative humidity in the green house.
Harvest and measurement: The radish plants were harvested upon attaining marketable maturity. The plants were dug out with their root system and washed thoroughly with deionised water, then placed on tissue paper to remove excess water. After removing the water, the tops and roots were separated and fresh weight was recorded for each treatment pot. The separated parts were then dried in aluminium dishes at 80°C for 48 h in a large oven and dry weight was recorded for each treatment pot.
Plant analysis: After oven drying, the plant samples were ground using a Tema mill which was cleaned thoroughly with a brush and acetone for each treatment and analyzed for Total-N by C/N analyzer the method as described by Watkins and Barraclough (1996). While P, S and B content by dry ashing technique as suggested by Isaac and Kerber (1971) and using ICP-OES for elemental analysis. Multi-element standard solutions were prepared of low, moderate and high concentrations in the same matrix as the samples for each element to calibrate the ICP-OES before introducing the samples and the results printed on Dec-writer II input/output terminal.
Statistical analysis: Statistical analysis of all the data collected during investigations were performed by MSTAT-C computer package and the means were compared by the LSD-test of significance (Steel and Torrie, 1980).
RESULTS AND DISCUSSION
Fresh matter yield: Fresh matter yield is an important component of radish crop and vegetable growers are always interested in its quantity as well as quality. The results showed that maximum yield of both tops and roots were obtained from the treatment receiving 0.5 mg B L-1 followed by 0.25 mg B L-1 (Fig. 1), but no significant difference was found between the yields of these two treatments, indicating a narrow range (as expected) for B sufficiency. A similar non significant difference was found by Scripture and McHargue (1945) between 0.25 and 0.5 mg B L-1 in sand culture study. The treatment received 0.5 mg B L-1 seems to be an optimum level of soluble B for radish, which showed the best growth response. These findings are supported by the previous work of Coetzer et al. (1990) and Buzetti et al. (1990). Such treatment showed 58.28% increase for tops and 65.6% increase for roots over control, however, beyond the 0.5 mg B L-1 the yield is significantly decreased due to toxic levels of added B. This is good evidence for demonstrating that the radish crop responds to added B in a limited range, above which toxic levels are reached causing a subsequent decline in yields. The conclusion can be drawn from the overall results that almost similar trends exist both in tops and roots with regard to fresh yield. However, the roots fresh weight was considerably more than tops, which in fact the storage organ of radishes contains high watery juice and are succulent in the fresh state. The analysis show that each unit increase in B supply above 0.5 mg B L-1 reduced the fresh matter yield of radish, suggesting excess B supply reduced the size of bulbs and damaged the leaves of plants.
Dry matter yield: Dry matter yield is an important criteria for the estimation of nutrient content and total uptake by plants. Dry matter yield of tops and roots were recorded separately, when fresh plant materials were oven dried until a constant weight was established. Results showed that again, the maximum dry matter yield of tops and roots were obtained from the treatment receiving 0.5 mg B L-1 (Fig. 1) but the yield of this treatment was found statistically at par with 0.25 mg B L-1 as in the case of with fresh matter yield.
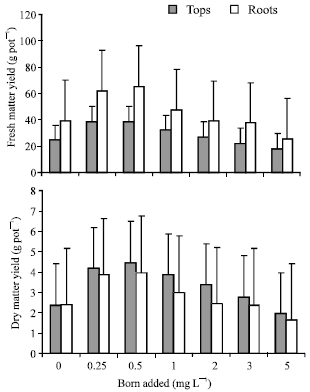 | |
| Fig. 1: | Influence of added boron on the yields of radish plants |
This treatment also showed 88.89% increase for tops and 66.11% increase for roots over control. This indicates that the radish crop responds well within the normal range of added B. Similar results were reported by Agarwala et al. (1978) and Orlova et al. (1980). Results also revealed that further increased in B supply causes a subsequent decline in the yields. These decrease in yields are mainly due to the toxic effects of B, resulting in reduced size and weight of plants. These results also indicate that radishes are more sensitive to B toxicity than deficiency under the conditions of the experiment. Dry matter yield of tops and roots decreases in a similar way as in the case of fresh matter yield, but there is difference found in the weight of roots. In fresh matter yield the weight of roots were considerably more than tops, while in dry matter yield the roots are lower than tops and this is due to a difference in the moisture content of roots and leaves.
Anions concentration and total uptake
Nitrogen: Results showed that the addition of B has no significant effect on the concentration of N in plants (Table 2). Normal levels of added B resulted in slightly lower concentrations in tops, but in roots remained constant in all treatments.
| Table 2: | Influence of added boron on the anions concentration in radish plants |
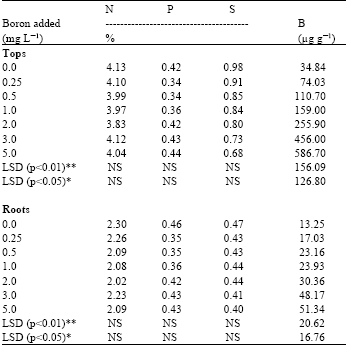 | |
| *, ** = indicate significance at p<0.05 and p<0.01 levels, respectively, NS = Non Significant | |
Results revealed that B deficient and toxic levels yielded slightly higher percentages of total N, which suggests that after stoppage of growth, the absorption of N continued in the B deficient and toxic plants. However, on the basis of these results and in agreement with the literature (Bonilla et al., 1980; Shelp and Shattuck, 1987; Willett et al., 1985) it seems that B must be functional in the metabolism and translocation of carbohydrates. Whether this function is direct or indirect through its effect on N metabolism can not be determined under the conditions of the experiment. On the other hand, B has significant treatment effects on the total uptake of N by both plant tops and roots (Table 3). Results also show that low uptake of N occurred at the 0 and 5 mg L-1 levels of added B. This indicates that extreme deficient or excess levels of B causes low uptake by plants. Moreover, comparing the data trend with growth response, the data indicates that the N and dry matter yields were similarly affected by varying levels of added B.
Phosphorus: Results revealed that although non-significant differences were found in the P concentration of tops and roots of plant (Table 2), but there is an up ward trend with increasing B in the nutrient solution. An enhancement in P content with higher levels of B was also observed by Pal et al. (1989).
| Table 3: | Influence of added boron on the total uptake of anions by radish plants |
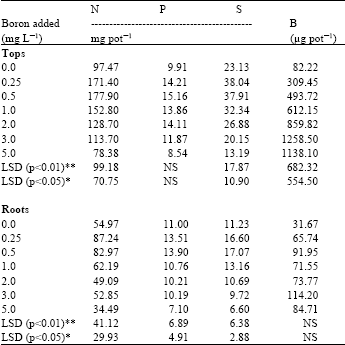 | |
| *, ** = indicate significance at p<0.05 and p<0.01 levels, respectively, NS = Non Significant | |
It is evident from the results that reduced P concentration at normal levels of added B seems to be due to the dilution effect, as the plant growth increased the demand for P was also increased. Similar, results were reported by Garate et al. (1984) and Singh et al. (1990). The data also indicate that almost equal P concentrations were found in both plant parts of the plant. So, it seems that P was equally distributed in plant tops and roots during the growth period. However, literature regarding the effect of B on the behavior of N and P is scarce and no mechanism has been proposed for B-P relationships other than that B and P have similar reactions with OH-1 groups, thus the uptake of these elements by plants is likely to follow similar pattern (Bartlett and Picarelli, 1973). The proposed mechanism can only be linked in the case of P concentration in plants at higher levels of added B. Results show a significant treatment effect of B on the uptake of P (Table 3), indicating that B levels influenced the P uptake in the same way that it did to the dry matter yield. Results further shows that the P uptake was increased at 0.5 mg B L-1 and decreased when no B was added or with highest level of added B. Low P uptake at deficient and toxic levels of added B may be due to functional relationships between these nutrients in plants. Kabata-Pendias and Pendias (1984) reported that the biochemical functions of B e.g., carbohydrate metabolism and transport of sugars through membranes, nucleic acids (DNA and RNA) synthesis, tissue development and formation of cell walls. All these functions are closely related to P. Therefore, it can be assumed that the borate ions may be fixed in the cell wall as an organic complex and would be responsible for a change in the permeability of the cell wall, resulting in a reduction of phosphate uptake and carbohydrates, under the deficient and toxic conditions of added B. Similar conclusions were drawn by Robertson and Loughman (1974).
Sulpher: Results show that with increasing B in the nutrient solution the concentration of S in tops and roots decreases (Table 2). These findings are in agreement with an early work of Parks et al. (1944) and later on confirmed by Valmis and Ulrich (1971). These investigators reported that with increasing B supply in the nutrient solution the concentration of S significantly decreases in tomato leaflets and in sugar beet mature blades, respectively. However, in the present study, differences found in the S concentration among the treatments are statistically non significant. Results indicate as the B toxicity in the substrate increases the concentration of S decreases, perhaps due to the competition of borate and sulphate ions on the root cells, which impaired the rate of S absorption by roots. On the other hand, with increasing B supply the total uptake of S significantly decreases in plant tops as well as in roots (Table 3) and run parallel to dry matter yield, suggested both were affected by applied B in the same manner, which showed similarity in the data. However, the decreasing trend of the S uptake due to B toxicity did not show a clear picture, whether this was the direct effect of B or due to some other ions like P was involved which affected the uptake of S by plants. This need further investigations.
Boron: B concentration in plants showed a significantly linear and positive relationship between B in tops (r = 0.98) and roots (r = 0.92) and B in nutrient solution (Table 2). This indicated that the accumulation of B in the leaves of plants depends only on the B levels in the root media (Salinas et al., 1986). Results revealed that the plant tops contain a higher concentration of B than roots, suggesting translocation through the xylem stream and transpiration involved in the accumulation of B in leaves. Shelp et al. (1987) and Oertli and Richardson (1970) have also emphasized that leaf venation, xylem stream and transpiration as factors primarily involved in the accumulation of B in leaves. Results also showed the B uptake by plants increases with each increment of added B except the treatment received 5 mg B L-1 (Table 3). Results indicate that the reduction in B uptake at highest B level is due to either a growth response or due to the lower or higher concentrations of other elements in the respective treatment, created unbalanced nutrient ratios. These results are however, in line with the early work of McIlrath et al. (1960). It is also evident from the results that more uptake was obtained in the tops than roots as in the case with B concentration in plants. These results agreed with the previous findings of Kluge (1990) who reported that total uptake was concentrated more in the aerial part, whereas in the root less accumulated, suggesting passive acropetal transport of B with the transpiration stream. It is well understood that the range between deficiency and toxicity of B is very small which is also observed in the present study for radish plants. In the present study good growth was found at B concentration 74 to 159 μg B g-1 DM in the tops and 23 to 24 μg B g-1 in the roots. Toxic concentrations were possibly attained, when leaves and roots contained 256 to 586 and 48 to 51 μg B g-1 DM, respectively. Gupta (1983) also listed deficient, sufficient and toxic levels for radish (cv. Cherry belle) tops when the roots began to swell as < 9, 96-217 and > 217 μg B g-1 DM. However, comparing the present results with Gupta’s results, the differences found are likely due to the different cultivar of radish used.
CONCLUSIONS
The following conclusions were drawn from the present sand culture study.
| • | Significant treatment effects were found on the growth response of radish plants and maximum fresh and dry matter yields were recorded at 0.5 mg L-1 of added B. Toxic effects accompanied by considerable yields decreases were observed at higher levels of B supply. |
| • | The concentration of P and B in plants were increased, S decreased and N remain unchanged. While, the total uptake of all anions except B decreased with increasing levels of B in the nutrient solution and showed close similarity to the growth response of radish plants, suggesting that total uptake of anions was useful but alternative way of estimating the nutrient behavior with regard to B supply. |
| • | The relative amount of all anions were higher in tops compared to roots. The present study suggests that the deficiency or excess of B affects the relative values of individual anions. Generally, low and high levels of added B had detrimental effects on the concentration and total uptake of anions. |
REFERENCES
- Agarwala, S.C., S. Farooq and C.P. Sharma, 1978. Interaction of boron supply and nitrogen source on growth, boron uptake, nitrogen fractions and sugars in radish. N' assimilation and crop productivity. Proceedings of the National Symposium Hissar, October 1978, New Delhi, India, Associated Publishing Company, pp: 185-194.
- Bartlett, R.J. and C.J. Picarelli, 1973. Availability of boron and phosphorus as affected by liming an acid potato soil. Soil Sci., 116: 77-83.
Direct Link - Bonilla, I., O. Cadahia, O. Carpena and V. Hernando, 1980. Effects of boron on nitrogen metabolism and sugar levels of sugar beet. Plant Soil, 57: 3-9.
CrossRefDirect Link - Gupta, U.C. and J.B. Sanderson, 1993. Effect of sulfur, calcium and boron on tissue nutrient concentration and potato yield. J. Plant Nutr., 16: 1013-1023.
Direct Link - McIlrath, W.J., J.A. Debruyn and J. Skok, 1960. Influence of boron supply on the micronutrients content of setaria shoots. Soil Sci., 89: 117-121.
Direct Link - Parks, R.Q., C.B. Lyon and S.L. Hood, 1944. Some effects of boron supply on the chemical composition of tomato leaflets. Plant Physiol., 19: 404-419.
PubMed - Pollard, A.S., A.J. Parr and B.C. Loughman, 1977. Boron in relation to membrane function in higher plants. J. Exp. Bot., 28: 831-841.
Direct Link - Robertson, G.A. and B.C. Loughman, 1974. Reversible effects of boron on the absorption and incorporation of phosphate in vicia faba L. New Phytol., 73: 291-298.
Direct Link - Salinas, R., A. Cerda and V. Martinez, 1986. The interactive effects of boron and macronutrients (P, K, Ca and Mg) on pod yield and chemical composition of Pea (Pisum sativum). J. Hortic. Sci., 61: 343-347.
Direct Link - Shelp, B.J. and V.I. Shattuck, 1987. Boron nutrition and mobility and its relation to the elemental composition of greenhouse grown root crops: Rutabaga. Commun. Soil Sci. Plant Anal., 18: 187-201.
Direct Link - Singh, J.P., D.J. Dahiya and R.P. Narwal, 1990. Boron uptake and toxicity in wheat in relation to zinc supply. Fertil. Res., 24: 105-110.
CrossRef - Watkins, N. and D. Barraclough, 1996. Gross rates of N mineralization associated with the composition of plant residue. Soil Bio. Biochem., 28: 169-175.
CrossRef - Willett, I.R., P. Jakobsen and B.A. Zarcinas, 1985. Nitrogen-induced boron deficiency in lucerne. Plant Soil., 86: 443-446.
CrossRef