
P.N. Rapando
Not Available
A.W. Wangai
Not Available
I.M. Tabu
Not Available
J. Ombiri
Not Available
R. Ramkat
Not Available
Asian Journal of Plant Sciences
Year: 2006 | Volume: 5 | Issue: 2 | Page No.: 311-315







ABSTRACT
Tomato spotted wilt topspovirus (TSWV) disease was recently reported to be endemic and posing a threat to cucumber production in Nakuru District, Kenya. A study was conducted to determine the response of commercially available cucumber varieties (Ashley, Marketer, Super Marketer and Chinese) to the disease and the effect of time of onset on disease severity and crop yield. The study was carried out in a green house where the four varieties were inoculated at cotyledon, 3-4 leaf stage and flower bud stage of growth. The experiment was arranged in a split plot design replicated four times. Variety Marketer was more tolerant to the disease compared to other varieties. All the varieties were generally tolerant to the disease at the cotyledon stage at the 3-4-leaf stage and the flower bud stage the plants were however susceptible to the disease. It was therefore concluded that planting variety Marketer and controlling the disease especially at 3-4 leaf and flower bud stages could form important components of integrated pest management programme of TSWV.
PDF Abstract XML References Citation
How to cite this article
P.N. Rapando, A.W. Wangai, I.M. Tabu, J. Ombiri and R. Ramkat, 2006. Effect of Inoculation on Severity of Tomato Spotted Wilt Disease in Cucumber. Asian Journal of Plant Sciences, 5: 311-315.
DOI: 10.3923/ajps.2006.311.315
URL: https://scialert.net/abstract/?doi=ajps.2006.311.315
DOI: 10.3923/ajps.2006.311.315
URL: https://scialert.net/abstract/?doi=ajps.2006.311.315
INTRODUCTION
Cucumber (Cucumis sativus) valued for its nutritional and cosmetic purposes is one of the important horticultural crops whose yield and quality has been declining because of diseases, like powdery mildew, downey mildew, cucumber mosaic and Tomato spotted wilt tospovirus (TSWV) (Wilson, 1999; Wetering et al., 1999). Yield losses between 40 to 50% in cucumber have been attributed to viral diseases (Helter et al., 1995). Tospoviruses belonging to the family Bunyaviridae are a group of emerging plant viruses transmitted by different species of thrips (Roggero et al., 1999). The most important viruses in the genus tospovirus include Tomato spotted wilt virus (TSWV) and Impatiens necrotic spot virus (INSV) (Ullman et al., 1998). In Kenya, TSWV was first reported on tomatoes in Nakuru district (Wangai et al., 2001). Preliminary research shows that cucumber is also susceptible (Wangai et al., 2001). The disease can cause economic losses as high as 100% in tomatoes and 60% in lettuce (Wangai et al., 2001; Pappu, 1997).
The underlying principle in the management of TSWV lies in the control of the primary vector, the Western flower thrips (Frankliniella occidentalis Pergande), which is predominant in Nakuru region (Soria and Mollema, 1995; Lelgut et al., 2001). Conventional control of this pest using pesticides is difficult because of its secretive habits within flowers and buds that prevent penetration hence limiting control (Seaton et al., 1997). Continuous use of pesticides also pollutes the environment and is deleterious to beneficial organisms not to mention that some strains of the pest are resistant to pesticides (Seaton et al., 1997; Jarosik et al., 1997; Hatalala et al., 1999). The wide host range of TSWV including several weed species implies that control can be achieved by combining several methods (Boiteux, 1995). Integrated pest management approach remains the best approach as it combines the utilization of chemicals with other control methods (Altieri, 1998). A combination of host plant resistance and reflective mulches has been found to significantly reduce TSWV on tomato (Riley and Pappu, 2000). An experiment was therefore carried out to determine the effect of genotype and stage of inoculation on TSWV severity and yield of cucumber.
MATERIALS AND METHODS
Study site: The experiment was carried out from June to December 2004 at National Plant Breeding Research Centre (NPBRC)-Njoro Nakuru District Kenya which lies at an altitude of 2160 m, longitude 36° East and latitude 0°20’. The soils are deep, well drained, dark to very dark reddish friable clay loams classified as vitric Mollic andosols (Jaetzold and Schmidt, 1983). The area receives about 1600 mm of rainfall per annum and temperatures ranging from 8-10.5 and 11.5-23°C.
Test plants: Cucumber seedlings were pre germinated on germination boxes and transplanted into plastic pots (12.5 cm diameter) containing steam-sterilized mixture of forest topsoil and gravel chips at a ratio of 9:1. Di-ammonium phosphate fertilizer (18% N: 46%:P2O5) at the rate of 1kg of fertilizer for 250 kg of the soil mixture was thoroughly mixed and used for growing seedlings in an insect free glasshouse. As an added precaution the green house was sprayed on a biweekly basis with Mitigan®(mites control) at a rate of 20 mL/30 L of water and metasystoc® to control aphids, whiteflies and thrips at a rate of 20 mL/45 L of water.
Inoculation of test plants: Leaves of tomato plants showing symptoms of TSWV were collected from a farmer’s plot in Njoro, Division, Kenya and crushed in a mortar using an extraction buffer at ratio (buffer to the volume of leaves) of 1:2. Test plants were pre darkened for 24 h prior to inoculation, to make them more susceptible to viral infection. Before inoculation, plants were dusted with carborandum of mesh 200, to act as an abrasive during inoculation. The extracted inoculum was then rubbed on the young leaves. Health controls were dusted with carborandum and rubbed with buffer minus the inoculum. The dusted plants were washed with distilled water and then transferred back to the dark room to increase the rate of viral infection. The plants were then transferred back to the green house the next morning.
Disease evaluation: Infection was determined by visual observation of symptoms on the leaves and confirmed by serological tests. Enzyme linked immuno-sorbent assay (ELISA) procedure (CABI/EPPO, 2002) was used for diagnosis and detection of TSWV in plant samples. The antibodies used were obtained from AgDia ELISA kit (Golnaraghi et al., 2001). The symptoms were observed 4-7 days after inoculation. The intensity of symptom expression was then scored on a scale of 0-5 i.e., 0 indicates lack of symptoms while 5 indicates a dead test plant after being severely affected (Dietz et al., 2001).
Treatments: Seeds of four cultivars (Marketer, Ashley, Chinese and Super-Marketer) were pre-germinated for two weeks at room temperature, then selected for uniformity and transplanted into individual plastic pots of 13 cm diameter and 20 cm long. At cotyledon, 3-4 and flower bud stages of growth, plants were inoculated with the viral inoculum. Healthy controls were not inoculated with the virus. The experiment was laid out in a split-plot design replicated four times. Stages of inoculation were the main plots while the varieties were the sub plots. Ridomil® (Metaxyl Mancozeb) and Metasystoc® (Oxid emeton methyl) at a rate of 2.5 kg ha-1 were used to control fungi and insect pests, respectively whenever necessary. The plants were randomly arranged in the green house in four blocks according to inoculation dates with each inoculation time randomized separately.
RESULTS
Disease symptoms: The disease symptoms included necrotic lesions, extensive chlorosis, systemic necrosis, severe stunting and sometimes death of plants. These typical symptoms of TSWV have been observed in potatoes and soybeans (Golnaraghi et al., 2001; Pourrahim et al., 2001). The symptoms however differed with variety. After inoculation, Ashley and Chinese varieties developed venal chlorosis followed by necrotic spots, which finally resulted in necrotic ring spots. Similar symptoms have been observed in squash (Cucurbita eppo) following infection with TSWV (Pappu et al., 1998). After necrosis, inoculated leaves abscised limiting the systemic movement of virus and leading to reduction of the amount of the virus in the plant (Torre et al., 2002). Towards plant maturity apical/bud necrosis was observed in infected plants. This is a common symptom observed following TSWV infection in groundnut (Salomone et al., 2003). On fruits, symptoms included malformation, yellowing, premature abortion and production of spongy, small and light fruits.
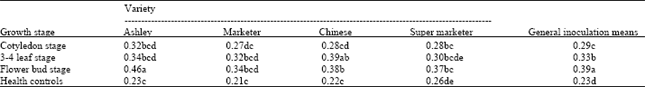
Disease rating: Generally the 3-4 leaf stage had the highest disease ratings followed by flower bud and cotyledon stages (Table 1).
This implies that 3-4 leaf stage was most susceptible while cotyledon was the least. Disease ratings were high on all inoculated plants of the four varieties. Apart from the flower bud stage where Chinese was high, Ashley was the most susceptible followed by Chinese variety and Marketer and lastly super Marketer. The disease rating was significantly related to the ELISA values (Table 2).
Yield and yield components
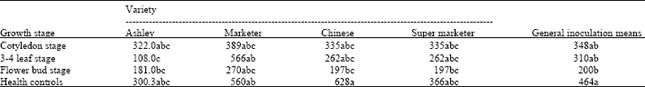
Fruit weight (g/plant): In absence of disease, Chinese variety had the heaviest fruits followed by Marketer, Ashley and lastly Super Marketer (Table 3). The effect of TSWV on cucumber varied with variety and stage of inoculation. The disease caused a 57 and 32% yield reduction in Chinese and Ashley compared to a 27% reduction in Marketer and Super Marketer. Performance of individual varieties differed with stage of inoculation. Inoculation of Ashley at the 3-4 and flower bud stages resulted in the lowest fruit weight i.e., a reduction of 64 and 40%, respectively. Inoculation at the cotyledon stage did not severely affect fruit weights of Ashley.
| Table 1: | Effect of TSWV inoculation at different growth stage on disease ratings of cucumber |
 | |
| *Means followed by the same letter (s) a column are not significantly different (p≤0.05) | |
| Table 2: | Effect of TSWV inoculation at different growth stage on ELISA values of cucumber |
 | |
| *Means followed by the same letter(s) a column are not significantly different (p≤0.05) | |
| Table 3: | Effect of TSWV inoculation at different growth stage on fruit weights (g) of cucumber |
 | |
| *Means followed by the same letter (s) a column are not significantly different (p≤0.05) | |
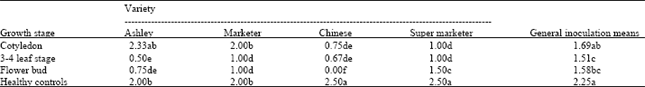
| Table 4: | Effect of TSWV inoculation at different growth stage on the number of marketable fruits of cucumber |
 | |
| *Means followed by the same letter (s) in a column are not significantly different (p≤0.05) | |
On contrary Marketer remained unaffected at the 3-4 leaf stage of inoculation but was severely affected at the cotyledon (31%) and flower bud stage (52%). Chinese variety was severely affected at the flower bud stage (68% reduction) followed by 3-4 leaf stage (58%) and cotyledon stage (58%). Super Marketer was only affected by the disease following inoculation at the flower bud stage where a 47% reduction in yield was observed. It is evident that all the varieties except for Marketer were severely affected following inoculation at the 3-4 leaf and flower bud stages.
Marketable yield: Chinese and Super Marketer had the highest number of marketable fruits followed by Ashley and Marketer (Table 4). Tomato spotted wilt virus (TSWV) disease caused a severe decrease in the marketable yield of the fruits.
A 71% reduction in marketable yield was observed in Chinese variety followed by Super Marketer, Ashley and Marketer with 54, 41 and 35% reduction, respectively. Generally marketable fruits were lowest after plants were inoculated at the flower bud stage followed by the 3-4-leaf stage and then cotyledon stages of inoculation.
Variety Ashley’s and Marketer’s marketable yield were severely reduced by TSWV at the 3-4 leaf stage and flower bud stage of growth while those inoculated at the cotyledon stage were not affected. The disease significantly reduced marketable yield of the Chinese variety and Super Marketer at all the stages of inoculation. All fruits of the Chinese variety were not marketable after plants were inoculated at the flower bud stage of growth.
DISCUSSION
The positive correlation of R2 = 0.4, between disease rating and serological (ELISA) test confirmed that most of the symptoms, observed were due to the virus. The reaction exhibited by the cucumber (necrotic spots and lesions that led to entire necrosis and death of plants in some cases) is referred to as hypersensitivity. Hypersensitive reaction has been noted to be common on cucumber plants expressing systemic acquired resistance to infection (Hammerschmidt 1999; Thomas and Jones, 2003). Paradoxically, this reaction is independent of the growth stage (Roggero et al., 1999). Necrotic lesions which were common on Ashley and Chinese and could have contributed to the high disease scores have also been observed in pepper, tomatoes and potatoes following mechanical inoculation (Golnaraghi et al., 2001). The Chinese variety lacked mature plant resistance because it took long to recover. Mature plant resistance was observed after inoculation of variety Ashley at the flower bud stage of growth as the plants had the least scores at this stage of growth. This type of resistance has been observed to be as a result of formation of tough tissues that do not allow wounding (Fraser, 1987).
The 3-4 leaf and flower bud stages were consistently susceptible. The high disease ratings were probably because the rapid growth at this stage may have initiated faster systemic movement of the virus to the actively growing plant parts. The mean ratings at the cotyledon stage of inoculation were lower than 3-4 leaf stage probably because of plant recovery at early stage of inoculation. Other research has also shown plant recovery after inoculation at the early stages of growth (Mathews, 1991). Paradoxically, other studies have found that transmission of TSWV is most effective at the cotyledon stage (Grovers et al., 1998).
There was a reduction in the yields both in quality and quantity after inoculation with the disease. Reduction in yields in terms of quality and quantity in tomatoes and pepper is a common effect of TSWV (Wangai et al., 2001; Riley and Pappu, 2000). Reduced yields of pepper that were significantly correlated with early infection of plants have also been observed (Gitaitis et al., 1998). The disease severely reduced fruit weights after plants were inoculated at the flower bud stage of growth. This could probably be due to the fact that the virus targeted the reproduction cells of the plants hence affecting cell division and multiplication and subsequently affected the yield. Viruses have similarly been noted to affect cell division leading to cell death, which interfere with physiological process in the plants (Bendahmane et al., 1999). Chinese variety was very susceptible to the disease and this was reflected on the weight and marketability of the fruits. The inoculated plants of this variety produced fruits that were yellowed and spongy leading to a reduction in fruit weights. The reason for the severe effect at the 3-4 leaf and flower bud stage was probably because of active plant growth that led to rapid movement and multiplication of the virus that ended up affecting fruit formation process. Variety Marketer remained unaffected at the 3-4 leaf stage probably because the variety could have limited movement of the virus as most plants at this stage of growth were stunted compared to other stages of growth.
Reduction in fruit quality/marketable yield out of viral infection has been observed in pineapple after infection with pineapple mealy bug wilt associated virus which caused non uniform fruit ripening and fruit malformation (Sipes et al., 2002). When plants are affected by TSWV, marketable yield is lowered due to systematic necrosis, which is severe on the fruits produced late in the season (Mavric and Ravnikar, 2001). On the other hand reduction in marketable yield of banana after infection with banana streak virus has been attributed to production of small and light fruits that are always malformed after the virus interferes with fruit development process (Daniellis et al., 2001).
There was some level of tolerance shown to TSWV on variety Marketer and Ashley at the cotyledon stage of growth as the scores were high but the yield was not affected. Variety Marketer was least affected by the disease at all the stages of inoculation. Variety Ashley can be rendered susceptible when the disease occurs at the 3-4 leaf stage. Chinese variety, though a high yielder in the absence of the disease, was severely affected by TSWV disease in terms of growth and yield of the crop irrespective of the stage of growth when the plants were inoculated. Generally, plants that were inoculated at the 3-4 leaf stage were the most susceptible. Moderate susceptibility was observed at the flower bud stage of growth and the cotyledon stage of growth was the most tolerant. A comparison between disease and yield variables showed Ashley to have some level of susceptibility, whereas Marketer was tolerant. Moderate susceptibility was observed on Super Marketer, as Chinese variety proved to be the most susceptible.
Analysis of the healthy plants show that Chinese and Marketer varieties had the highest yield followed by Super Marketer and lastly Ashley. The TSWV disease however varied with variety and inoculation stage. Although Chinese variety had a high yield potential, it suffered the highest yield loss as a result of the disease. Marketer and Super Marketer had the least yield reduction attributed to the disease. Stage on inoculation significantly affected the disease severity. At 3-4 leaf and flower bud stages, varieties were most susceptible to TSWV. Hence choice of variety and stage of growth could form important components of integrated pest management.
ACKNOWLEDGMENT
The authors are greatly indebted to Kenya Agricultural Research Institute for funding the study and allowing them to use their facilities.
REFERENCES
- Bendahmane, M., M. Koo, E. Karrer and R.M. Beachy, 1999. Display of epitopes on the surface of tobacco mosaic virus: Impact of charge and isoelectric point of the epitope on virus host. J. Mol. Biol., 290: 9-20.
PubMedDirect Link - Boiteux, L.S., 1995. Allellic relationships between genes for resistance to tomato spotted wilt tospovirus in Capsicum chinese. Theor. Applied Genet., 90: 146-149.
Direct Link - Daniellis J.W., A.D. Greering, N.J. Bryde and E.J. Thomas, 2001. Effect of banana streak virus on the growth and yield of dessert bananas in tropical Australia. Ann. Applied. Biol., 98: 23-27.
Direct Link - Dietz, M.J., F. Nuez, B. Ricarte and S. Rosseloo, 2001. Resistance to Tomato spotted wilt virus Resistance to Tomato spotted wilt virus introgressed from Lycopersicon peruvian in line UPV. Euphytica, 119: 357-367.
Direct Link - Gitaitis, R.D., C.C. Dowler and R.B. Chalfant, 1998. Epidemiology of Tomato spotted wilt in pepper and tomato in Southern Georgia. Plant Dis., 82: 752-756.
Direct Link - Hammerschmidt, R., 1999. Induced disease resistance: How do induced plants stop pathogens? Physiol. Mol. Plant Pathol., 55: 77-84.
CrossRef - Helter, N.B., J. Patricia and Y.M. Rodney, 1995. Control of Western flower thrips (F. occidentalis) pupae in compost. Ann Applied Biol., 127: 405-412.
Direct Link - Jarosik, V., M. Kolias, L. Lapchin, J. Rochat and A.F.G. Dixon, 1997. Seasonal trends in the population increase of Frankliniella occidentalis (Thysanoptera: Thripidae) on cucumber. Bull. Entomological Res., 87: 487-498.
Direct Link - Pappu, H.R., 1997. Managing tospoviruses through biotechnology progress and prospects. Biotechnol. Dev. Monitor, 32: 14-17.
Direct Link - Riley, D.G. and H.R. Pappu, 2000. Evaluation of tactics for management of thrips vectored Tomato spotted wilt virus in tomato. Plant Dis. J., 84: 847-852.
Direct Link - Sipes, B.S., D.M. Sether and J.S. Hu, 2002. Interaction between Rotylenchulus reniformis and Pineapple mearly wilt associated virus-1 in Pineapple. Plant Dis., 86: 933-938.
CrossRefDirect Link - Thomas, C.M.R. and A.C. Jones, 2003. Selection, biological properties and fitness of resistance breaking strains of Tomato spotted wilt virus in pepper. Ann. Applied. Biol., 142: 235-243.
Direct Link - Wetering, F.V., K. Posthuma, B.R. Gold and S.R. Peter, 1999. Assessing the susceptibility of chrysanthemum cultivars to TSWV. Plant Pathol., 48: 693-699.
Direct Link - Wilson, C.R., 1999. The potential of reflective mulching in combination with insecticide sprays for control of aphid-borne viruses of iris and tulip in Tasmania. Ann Applied Biol., 134: 293-297.
Direct Link - Golnaraghi, A.R., N. Sharaeen, R. Pourrahim, S. Ghorbani and S. Farzadfar, 2001. First report of Tomato spotted wilt virus on soy bean in Iran. Plant Dis., 85: 1290-1290.
Direct Link - Mavric, I. and M. Ravnikar, 2001. First report of Tomato spotted wilt virus and impatients necrotic spot virus in Slovenia. Plant Dis., 85: 1288-1288.
Direct Link