
R. Selvi
Not Available
A.R. Muthiah
Not Available
N. Manivannan
Not Available
T.S. Raveendran
Not Available
A. Manickam
Not Available
R. Samiyappan
Not Available
Asian Journal of Plant Sciences
Year: 2006 | Volume: 5 | Issue: 2 | Page No.: 277-280







ABSTRACT
The present study was undertaken to identify RAPD marker associated with Mungbean Yellow Mosaic Virus (MYMV) resistance in mungbean (Vigna radiata (L.) Wilczek) cross ML 267 x CO 4. Bulked segregant analysis was employed to identify RAPD markers linked to MYMV resistant gene of ML 267. A total of 149 random decamers were surveyed for identification of polymorphic markers between the DNA bulks of resistant and susceptible F2 individuals and their parents. Approximately 94% of these produced DNA amplification in both parents and bulks. Forty one primers produced specific band for resistant parent which were absent in susceptible parent. Out of 41 random primers, three primers viz., OPT 16, OPS 7 and OPAK 19 produced specific fragments viz., OPT 16564, OPS 7900 and OPAK 19400, respectively in resistant parent and resistant bulk, which were absent in the susceptible parent and bulk. Amplification of individual DNA samples out of the bulk with putative markers, OPS 7900 only revealed polymorphism in all 8 resistant and 6 susceptible plant, indicating that the marker OPS 7900 was associated with MYMV resistance in ML 267.
PDF Abstract XML References Citation
How to cite this article
R. Selvi, A.R. Muthiah, N. Manivannan, T.S. Raveendran, A. Manickam and R. Samiyappan, 2006. Tagging of RAPD Marker for MYMV Resistance in Mungbean (Vigna radiata (L.) Wilczek). Asian Journal of Plant Sciences, 5: 277-280.
DOI: 10.3923/ajps.2006.277.280
URL: https://scialert.net/abstract/?doi=ajps.2006.277.280
DOI: 10.3923/ajps.2006.277.280
URL: https://scialert.net/abstract/?doi=ajps.2006.277.280
INTRODUCTION
Mungbean (Vigna radiata (L.) Wilczek) is one of the important grain legumes. It is an excellent source of easily digestible good quality protein with low flatulence. Mungbean plays an important role in alleviating the protein malnutrition in developing countries. The average yield of mungbean is 341 kg ha-1 in India despite its yield potential of more than 1.5 t ha-1. There are many factors responsible for low productivity ranging from plant ideotype to biotic and abiotic stresses. Among the various diseases, the Mungbean Yellow Mosaic Virus (MYMV) disease was given special attention because of its severity and ability to cause yield loss upto 85% (AVRDC, 1998).
MYMV disease can be controlled by chemical, cultural and genetic methods. Nevertheless, if host resistance alone is sufficient to check the disease, then it is to be preferred over other methods because it is the least expensive and has no adverse environmental effects. Therefore, the development of varieties by incorporation of MYMV resistant genes into commercially acceptable cultivars is an effective strategy to check the disease. Identification of resistant lines through conventional method by field screening is time consuming and it requires evaluation at ‘hot spot’ area. Many times the disease incidence at the testing site may not be to the desired level. The disease incidence is seasonal and cannot be created as and when desired by artificial means. Therefore, the indirect selection using molecular marker linked to MYMV resistant genes will be helpful in rapid identification of genotypes carrying these genes without subjecting them to MYMV screening. Further, the resistant genotypes can be identified in the seedling stage itself thus reducing the time required for the development of resistant varieties. This is most important in case of back cross breeding programme. Pattnaik and Kole (2002) identified protein marker to differentiate resistant and susceptible genotype to MYMV in mungbean. However, this technique yields very limited number of polymorphisms making proteins marker is not very powerful when compared to PCR-based markers. Thus, the present investigation was carried out with an objective to identify the association of resistance with Random Amplified Polymorphic DNA (RAPD) marker.
MATERIALS AND METHODS
The present investigation was carried out at Pulses Breeding Station, Center for Plant Breeding and Genetics, Tamil Nadu Agricultural University, Coimbatore, Tamil Nadu State, India and National Pulses Research Center, Vamban, Tamil Nadu State, India during the period from 1999 to 2002.
Evaluation for MYMV reaction: The infector row method consisting of sowing two test rows alternating with spreader rows of susceptible variety, CO 5 mungbean was adopted. The test materials were scored after 80% of the plants in spreader rows showed MYMV incidence. The following rating scale suggested by Singh et al. (1988) is adopted. The mean disease score was calculated as (disease rating and frequency) per total number of plants scored (Table 1).
Development of mapping population: The MYMV resistant parent ML 267 was crossed with susceptible parent CO 4 during Summer, 2000 at Pulses Breeding Station, Tamil Nadu Agricultural University, Coimbatore. The crossed seeds were sown during Kharif, 2000 and true hybrid was fixed based on morphological character of F1. The true single F1 plant was selfed and forwarded to F2. The F2 seeds were sown during Summer, 2001. The 242 F2 individuals were tagged and leaf samples were collected and stored in -80°C for genotyping. Seeds from each F2 plants were collected separately and forwarded to F3 generation for evaluation of MYMV reaction.
Phenotyping of mapping population: The F3 seeds (25 numbers) from each F2 plants were sown in two replications during Kharif, 2001 at National Pulses Research Center, Vamban along with susceptible check CO 5. The susceptible check was sown 10 days earlier to test materials. The individuals in each F3 family were scored for MYMV reaction using 1-9 scale rating (Singh et al., 1988) given for each plant. The average rating and replication mean rating were calculated for each family. Based on the replication mean score, the homogenous resistant and susceptible F2 plants were identified.
DNA extraction:DNA from 19 selected F2 individuals and their parents were extracted following the method of Doyle and Doyle (1987) with modification. From frozen leaves stored at -80°C, 2 g of leaves were completely homogenized with liquid nitrogen.
| Table 1: | MYMV disease reaction scale |
 | |
Extraction buffer (100 mM Tris HCl (pH 8.0), 20 mM EDTA, 1.4 M NaCl, 2% CTAB L-1) was added in 50 mL tubes filled with leaf powder to the volume of 15 mL and mixed well. The tubes were incubated at 65°C for 30 min with repeated shaking. Equal volume of chloroform: isoamylalcohol mix (24:1) was added and mixed thoroughly for 15 min followed by centrifugation at 4000 rpm for 30 min. Equal volume of isopropanol was added to the supernatant. DNA was hooked out after half an hour and washed in 70 ethanol and suspended in 500 μL of TE buffer (pH 8.0). The DNA was incubated with 10-15 μL (10 μg μL-1 concentrations) of RNase for 30 min. To this equal volume of chloroform: isoamylalcohol was added and centrifuged at 12,500 rpm for 10 min. Twice the volume of absolute ethanol and 1/10th volume of sodium acetate were added to the aqueous layer and incubated overnight. The content was centrifuged at 12,500 rpm for 15 min and supernatant was discarded. The pellet was washed with 70 ethanol and air dried. Then the pellet was dissolved in 500 μL of TE and stored at -20°C. The quality and quantity of DNA were checked through 0.8% agarose gel by electrophoresis. DNA concentration for PCR reaction was estimated by comparing the band intensity produced by the known dilution that gave good amplification.
RAPD analysis: Genomic DNA was used as template for PCR amplification as described by Williams et al. (1990). Amplification was carried out with 20 μL reaction volume containing 10 mM Tris HCl (pH 8.3), 50 mM KCl, 1.5 mM MgCl2, 0.001% gelatin, dATP, dCTP, dGTP and dTTPs each at 0.1 mM , 0.2 mM primer (OPERON Technologies Inc.), 25-30 ng of genomic DNA and 0.5 unit of Taq DNA polymerase. Amplification was performed in 0.2 mL thin walled PCR tubes using PTC-100TM Programmable Thermal Controller (MJ Research Inc.) programmed for 40 cycles. After initial denaturation for 2 min at 94°C, each cycle consisted of one minute at 94°C, one minute at 37°C and 2 min at 72°C. The 40 cycles were followed by seven min final extension at 72°C. PCR amplified products were subjected to electrophoresis on 1.5% agarose gel in 1X TBE buffer at 120 V for 4 h using Hoefer Super Submarine electrophoresis unit (Pharmacia Biotech, USA). The electronic image of ethidium bromide stained gels were captured using Alpha Imager system.
Tagging of MYMV resistant genes with RAPD markers: Bulked segregant analysis was employed to identify RAPD markers linked to MYMV resistant gene of ML 267. Segregating population (F2) of the cross combination ML 267 X CO4 was separated into homozygous resistant and susceptible based on F3 phenotyping. The parents, ML 267 and CO 4 were screened along with resistant and susceptible bulks constituted from 8 resistant and 6 susceptible individuals of F2 population respectively, using 149 random primers obtained from OPERON Technologies Inc. The linkage of the polymorphic marker is confirmed using phenotypic evaluation of the individuals of the segregating population from which the bulks were generated.
RESULTS AND DISCUSSION
Advances in cellular and molecular biology have provided new tools like marker aided selection, which increases the efficiency of breeding programmes especially disease resistance breeding. In conventional methods, selection of resistant genotype relied on field or glass house screening. It may also require the use of controlled environment, when the disease development is temperature sensitive as that of yellow mosaic disease. Hence, the screening process is complex and time consuming. But in molecular marker assisted selection, once a gene of interest is tagged with molecular marker, selection for that gene can be made based on the marker rather than the phenotype of individual which embodies the gene. Moreover selection can be performed in the seedling stage itself by molecular marker (Tanksley, 1983; Beckmann and Soller, 1986).
In the present investigation, an attempt was made to identify the RAPD marker associated with MYMV resistance, using bulked segregant analysis. One of the most time consuming requirements in marker development is the need to screen entire mapping populations with every primer has been reduced by bulked segregant analysis. The screening of contrasting bulks made from individuals of same phenotype of segregating population, suggests that testing the entire population is required only when polymorphisms between the bulks are detected. This result in a considerable saving of time particularly when used with PCR based technique such as RAPD (Williams et al., 1990). Bulked segregant analysis has been used to detect markers linked to many disease resistant genes including Uromyces appendiculatis resistance in common bean (Haley et al., 1993), leaf rust resistance in barely (Poulsen et al., 1995) and angular leaf spot in common bean (Nietsche et al., 2000). The RAPD markers prepared by the polymerase chain reaction are used widely for mapping genes, because RAPD markers are more rapidly and easily detectable than RFLP markers (Welsh and McClelland, 1990; Williams et al., 1990; Foolad et al., 1993). The RAPD marker had also been used effectively for tagging disease resistant (Hu et al., 1997; Alzate-Marin et al., 1999; Sartorato et al., 1999) and virus resistant genes (Ohmori et al., 1995, 1996; Stevens et al., 1995; Gmitter et al., 1996).
Tagging of MYMV resistant genes with RAPD markers: Bulked segregant analysis was employed to identify RAPD markers linked to MYMV resistant gene of ML 267. A total of 149 random decamers were surveyed for identification of polymorphic markers between the DNA bulks of resistant and susceptible F2 individuals and their parents. Approximately 94% of these produced DNA amplification in both parents and bulks. Forty one primers produced specific band for resistant parent which were absent in susceptible parent. Out of 41 random primers, three primers viz., OPT 16, OPS 7 and OPAK 19 produced specific fragments viz., OPT 16564, OPS 7900 and OPAK 19400, respectively in resistant parent and resistant bulk, which were absent in the susceptible parent and bulk. Amplification of individual DNA samples out of the bulk with putative markers, OPS 7900 only revealed polymorphism in all 8 resistant and 6 susceptible plant (Fig. 1), indicating that the marker OPS 7900 was associated with MYMV resistance in ML 267. However, one of the F2 individuals (plant number 5) in the susceptible bulk generated OPS 7900 marker. This may be due to misclassification of heterozygote with resistant gene as susceptible. This problem could be overcome by screening a large number of plants per progeny. This type of misclassification was also encountered by Poulsen et al. (1995), when employing the bulked segregant analysis to identify a RAPD marker linked to leaf rust resistance in barley.
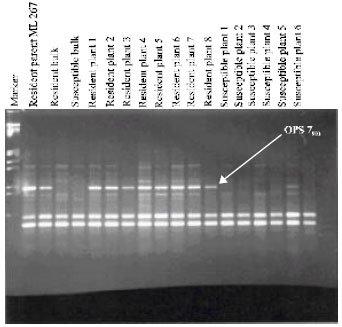 | |
| Fig. 1: | Cosegregation of RAPD marker OPS 7900 with MYMV resistance in F2 individuals of ML 267/CO 4 of mungbean |
The association of OPS 7900 RAPD marker with MYMV resistance can be employed for selection of MYMV resistant genotypes. However, problems associated with the reproducibility of the RAPD markers in different laboratories have been reported.
SCAR (Sequence Characterised Amplified Region) represent an alternative to increasing the reproducibility of RAPD markers (Adam-Blondon et al., 1994). Thus, the OPS 7900 should be converted into SCAR for increasing reproducibility and efficiency of marker for selection of MYMV resistant genotypes.
REFERENCES
- Adam-Blondon, A.F., M. Sevignac, H. Bannerot and M. Dron, 1994. SCAR, RAPD and RFLP markers linked to a dominant gene (Are) conferring resistance to anthracnose in common bean. Theor. Applied Genet., 88: 865-870.
CrossRef - Beckmann, J.S. and M. Soller, 1986. Restriction fragment length polymorphisms and genetic improvement of agricultural species. Euphytica, 35: 111-124.
CrossRefDirect Link - Doyle, J.J. and J.L. Doyle, 1987. A rapid DNA isolation procedure for small quantities of fresh leaf tissue. Phytochem. Bull., 19: 11-15.
Direct Link - Foolad, M.R., R.A. Jones and R.L. Rodriguez, 1993. RAPD markers for constructing intraspecific tomato genetic maps. Plant Cell Rep., 12: 293-297.
CrossRefDirect Link - Gmitter, Jr. F.G., S.Y. Xiao, S. Huang, X.L. Hu, S.M. Garnsey and Z. Deng, 1996. A localized linkage map of the citrus tristeza virus resistance gene region. Theor. Applied Genet., 92: 688-695.
CrossRefDirect Link - Ohmori, T., M. Murata and F. Motoyoshi, 1996. Molecular characterization of RAPD and SCAR markers linked to the Tm-1 locus in tomato. Theor. Applied Genet., 92: 151-156.
CrossRef - Poulsen, D.M.E., R.J. Henry, R.P. Johnston, J.A.G. Irwin and R.G. Rees, 1995. The use of bulk segregant analysis to identify a RAPD marker linked to leaf rust resistance in barley. Theor. Applied Genet., 91: 270-273.
CrossRefDirect Link - Tanksley, S.D., 1983. Molecular markers in plant breeding. Plant Mol. Biol. Rep., 1: 3-8.
CrossRefDirect Link - Welsh, J. and M. McClelland, 1990. Fingerprinting genomes using PCR with arbitrary primers. Nucleic Acids Res., 18: 7213-7218.
CrossRefPubMedDirect Link - Singh, G., S. Kapoor and K. Singh, 1988. Multiple disease resistance in mungbean with special emphasis on mungbean yellow mosaic virus. Proceedings of the 2nd International Symposium on Mungbean, Shanhua, Nov. 16-20, 1987, Asian Vegetable Research and Development Centre, Tainan, Taiwan, pp: 290-296.