
C.G. Ipsilandis
Not Available
B.N. Vafias
Not Available
A. Karagiozopoulou
Not Available
C.K. Goulas
Not Available
Asian Journal of Plant Sciences
Year: 2005 | Volume: 4 | Issue: 1 | Page No.: 75-82







ABSTRACT
Three F1 single-cross maize commercial hybrids were used in mechanical mixtures with their mother inbred parents aiming to explore the impact of the use of low-purity maize hybrid seed on leaf chlorophyll content and on yielding performance under varying plant densities. The evaluation of hybrid performance was conducted in two different experiments. Two plant densities were used in the first experiment and three in the second one. Starch gel electrophoresis was performed for hybrid seed purity determination. The seed emergence percentage (Emergence Index) in growth chamber was measured to estimate the differences between mixture seed partitions. In 11°C, the pure hybrid was easily distinguished from low purity seed partitions in the sixth day of measurement for all hybrids. Yielding performance in the field differed significantly for both factors: mainly between mechanical mixtures (seed partitions) and between a few seed partitions across densities. Leaf chlorophyll content was estimated on the basis of chlorophyll percentage (in SPAD units) in the leaves of the hybrid plants and their inbred parents. Differences were found mainly between hybrids and inbred lines and when seed partitions were compared within densities. In this study, it was demonstrated that seed purity is indeed a major stress in comparison to plant density and that an attentively conducted seed production program may insure yielding performance in the field for modern maize hybrids.
PDF Abstract XML References Citation
How to cite this article
C.G. Ipsilandis, B.N. Vafias, A. Karagiozopoulou and C.K. Goulas, 2005. F1 Single-cross Maize Hybrid Performance under Low Purity Conditions. Asian Journal of Plant Sciences, 4: 75-82.
DOI: 10.3923/ajps.2005.75.82
URL: https://scialert.net/abstract/?doi=ajps.2005.75.82
DOI: 10.3923/ajps.2005.75.82
URL: https://scialert.net/abstract/?doi=ajps.2005.75.82
INTRODUCTION
High and stable yielding performance is the main target for plant breeders of maize hybrids[1,2]. Higher grain yield productivity of modern maize hybrids resulted indirectly by improving a range of traits associated with tolerance to various biotic and abiotic stresses and by improving the efficiency of capture and use of resources[2-4]. Many researchers[3,5-10] mention the response of maize hybrids in several stress conditions. Tollenaar[9] concluded that the plant density might be a major stress to older hybrids with minor impact to newer ones. Also, the differential response to stress between older and newer hybrids has been shown for low night temperature during the grain-filling period by Dwyer and Tollenaar[6] and low soil moisture in the field[7]. Differences in grain yield between older and newer maize hybrids were shown to be a function of plant population density[11]. At lower plant densities, the differences between older and newer hybrids were small, becoming greater as plant density increased[3,9,12]. Vafias et al.[10] showed that the hybrid yield was reduced in the presence of the inbred line within the plots, especially in low plant density. High plant density favours the impure hybrid performance, when lower plant density favours the pure hybrid performance and the inbred line expression. Fasoula and Fasoula[13] distinguished three components of crop yield potential: the potential yield per plant in the absence of competition, the tolerance to stresses and the responsiveness to inputs. Tokatlidis[14] concluded that the higher potential yield per plant decreases the optimum plant density and renders the hybrids less density-dependent.
Hybrid seed purity is an important parameter that insures high yielding performance[1] and thus is of great consideration in seed production programs[15,16]. Craig[17] depicted careful plant and ear selection by trained technicians must be practiced throughout the growing season and at harvest to eliminate individual plants which exhibit phenotypes varying from the established accepted phenotype of the inbred. He also mentioned that isolation of seed fields and pollen control are extremely critical factors for the insurance of hybridisation of intended cross. Genetic purity of intended cross is dependent upon various parameters that must be controlled. Even more important, special care should be taken to insure a proper nick of seed parent silks and pollen from the pollinator. This study recommended seed purity and germination tests to ensure the physical quality of seeds. The basic reason for genetic impurity of hybrid seed is the presence of the mother’s inbred line seed in the seed partitions, as a result of incomplete emasculation[15,18]. Wych[18] mentioned that, this kind of impurity is usually present at a level of 3-5%, without being detected in the farmer’s field and without any effect on the yielding performance.
Starch gel electrophoresis and other techniques were used for seed purity determinations[16,19]. On the other hand, several physiological parameters were analyzed, in order to determine maize performance under variable conditions, mainly based on photosynthesis estimation[6,7,20]. Increased plant-to-plant uniformity for yield per plant of maize hybrids, with decreasing the plant density, have been found for traits like dry weight and plant size[20].
The purpose of this study was to explore the impact of low-purity maize hybrid seed usage on leaf chlorophyll content and on yielding performance under varying plant densities and compare the impurity stress to high population stress.
MATERIALS AND METHODS
Genetic materials: From three commercial single-cross maize hybrids (named Anthipi, Aris and Dias) seed partitions were formed by mixing mechanically the hybrid seed with the respective inbred seed of mother-parent. Two different groups of seed partitions were used in separate experiments held in the areas of Larissa and Karditsa, Greece. In the first group in the area of Larissa, five seed partitions were formed based in the percentage of the inbred parent (mother), present in the mechanical mixture. These seed partitions consisted of 0, 10, 20, 30 and 40% inbred line seed, in mixture with the pure hybrid seed. In the second group in the area of Karditsa, six seed partitions were formed based in the percentage of the inbred parent present in the mechanical mixture. These seed partitions consisted of 0, 10, 15, 20, 25 and 30% inbred line seed in mixture with the hybrid seed.
Field experiments and yield estimation: During 1999 growing season the materials were evaluated in the two separate experiments arranged in split-plot complete block designs, under three or two different plant densities (in Larissa and Karditsa, respectively) and four replications per density (with the subplots consisting of plant densities). The three plant densities in Larissa consisted of 47600, 63500 and 95200 plants ha-1 and the two densities in Karditsa consisted of 70000 and 84000 plants ha-1. The three different hybrids for each experiment was preferred to be evaluated in different but adjacent fields within each area (Larissa or Karditsa) to obtain more representative results. Plots consisted of two rows, 800 cm long and 75 cm wide. Border rows were not used in purpose, because of the need of grower’s field simulation (field effects). Distances between plants within rows were properly selected to obtain the desired plant densities. The general scheme was: 2 experiments (places) X 3 hybrid fields X 5 or 6 different seed partitions X 3 or 2 densities X 4 replications. For technical reasons due to lack of seed, hybrid Anthipi was sown in one density (63500 plants ha-1). The hybrid/inbred hills were planted randomly and plots were overplanted and thinned to desired stand at the seedling stage (4 young leaves). The separate inbred line hills were marked properly (by using coloured seed and then flags), in a way that the inbred plants were recognised easily after germination. Nitrogen and P fertiliser were applied at the rate of 120 and 60 kg ha-1 respectively at planting stage, while additional N (100 kg ha-1) was applied seven weeks after planting. Complete weed control was obtained by hand weeding. Trials were regularly irrigated to avoid drought stress. Plants were harvested individually and, after adjusting to 15.5% grain moisture, the yields per plot were calculated. Analysis of Variance, correlations and departure from linearity were based on common procedures[21].
Electrophoresis procedure: Starch gel electrophoresis was performed for seed purity determination. About the procedure, 25 specimens were used per gel plate. Every four hybrid seeds, one inbred parent seed was used as a check. In that way, for every gel, 20 hybrid seeds were used followed by 5 checks (inbred line). Only seeds of the same hybrid were used in a specific gel plate. In that way, three different gels were used for each one of the three hybrids. Four replications per gel were used, so 12 gels were formed in total (for all hybrids). Starch gel electrophoresis in four isozyme systems was conducted. The systems used were the non-specific esterase isozyme system (EST), the acid phosphatase (ACP), the glutamic-oxaloacetic transaninase (GOT) and the isocitrate dehydrogenase (IDH). These isozyme systems were chosen because of the variability they exhibit in the analysis procedure and were the subject of numerous studies of polymorphisms in maize[22,23]. Each gel was cut into four slices, in order to perform the analysis in all isozyme systems. Specimen and gel preparations, as well as dying and analysis method, were based on the findings of other researchers[22-24]. Starch gel electrophoresis showed that there were no great differences between isozymic patterns, in almost all isozymic systems. Hybrid Dias was a uniform material without variability. Hybrids Anthipi and Aris, showed variability about 4-5% for IDH. In general, authentic seed (blueprint) was considered satisfactory.
Leaf chlorophyll content: Leaf chlorophyll content was estimated on the basis of chlorophyll percentage in the leaves of the hybrid plants and their inbred parents at flowering stage. The Minolta SPAD 502 instrument was used to record the chlorophyll percentage. The measurements involved at least 7 hybrid and 3 inbred line plants in each plot. Four measurements per plant were conducted, involving the leaves near the spike.
The emergence percentage estimation: For the material used, the emergence percentage (Emergence Index) in growth chamber was used to estimate the differences between mixture seed partitions. Fifty seeds from every seed partition were evaluated for germination in a properly prepared box (seeds were set between paper sheets). Four replications (boxes) for every seed partition were used per hybrid. The seed partitions used, consisted of the hybrid seed contaminated by the inbred parent seed as was mentioned above for the material used in the area of Karditsa (0, 10, 15, 20, 25, 30% percentage of the inbred parent), inbred line seed being marked properly. Two different temperatures were used in the growth chamber: 11 and 15°C, respectively. The general scheme was: 2 temperatures X 3 hybrids X 6 different seed partitions X 4 replications (boxes) X 50 seeds/box. Tap water was used to moisture the seeds and fungicide (Benlate) was added. Measurements involving the number of germinating seeds were repeated every three days starting from the third day after the seeds were put for germination into the boxes and ending at the seventeenth day. The emergence percentage was calculated on the basis of Emergence Index (EI) according to the adjusted formula[25]:
Where:
| N | = | Number of plants germinated in one day (in an interval of three days) |
| D | = | Days after the first day (when seeds were put for germination into the boxes) |
| T | = | Total number of plants that were germinated at the seventeenth day |
RESULTS AND DISCUSSION
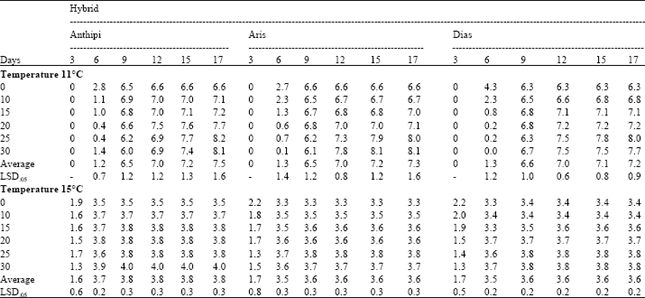
Laboratory procedure: The Emergence Index (EI) was found to be a satisfactory measurement to distinguish between different seed partitions (Table 1), after blue print specifications of original seed were insured by electrophoresis procedure. In 11°C, there were a few statistically significant differences, mainly for hybrid Dias. In the sixth day the different seed partitions of hybrid Dias, ranged from 0 to 4.3 (LSD0.5=1.2) and even in the last day the seed partitions ranged from 6.3 to 8 (LSD0.5=0.9). The pure hybrid germinated better than low purity seed partitions, since EI reached 4.3 in the sixth day and completed the germination in the ninth day. On the contrary, in 15°C, there were significant differences between seed partitions for each day of measurement (LSD values ranged from 0.2 to 0.8). Seeds in 15°C germinated faster in comparison to 11°C, so that germination was completed within six days, for almost all hybrids. In 11°C, germination was completed after about 15 days. In this temperature, the pure hybrid was easily distinguished from low purity seed partitions in the sixth day of measurement for all hybrids, since there were significant differences. In both temperatures pure hybrid Dias showed faster germination during the first few days and all the pure hybrids were always better germinating in comparison to low purity seed partitions (pure hybrid showed higher EI within first days and reached the maximum EI faster).
The yielding performance: Yielding performance (kg ha-1) of different genetic materials was influenced significantly by the seed purity conditions (Table 2). The seed partitions differed significantly for all hybrids and across experimental areas (different places). Differences between densities were of minor importance, since only hybrid Aris was influenced significantly by the stress of plant density in the area of Larissa (LSD0.5=890 kg ha-1). Hybrid Aris as an older hybrid than Anthipi and Dias is expected to exhibit a less stable yielding performance than newer ones. According to Tollenaar and Wu[4], Vafias et al.[10], Tokatlidis et al.[26], Ipsilandis and Vafias[27] modern hybrids show better and stable yielding performance than older ones under stress conditions. Table 2 shows that, hybrid Aris was unstable across densities (pure hybrid ranged from 11500 to 14080 kg ha-1 in Larissa) and was favored by the environment in the area of Larissa. It was found strong negative association between yield and seed purity (Fig. 1) for all hybrids in the two areas.
| Table 1: | Emergence Index (EI) values obtained every three days (3-17) in growth chamber, for three hybrids (Anthipi, Aris, Dias), in two temperatures (11 and 15°C), for the 6 seed partitions (0-30%) |
 | |
| Table 2: | Yielding performance (kg ha-1), for three hybrids (Anthipi, Aris, Dias), in two areas (Karditsa, Larissa), in the respective (6 or 5) seed partitions (part) and plant densities (dens), with average values (Avg) |
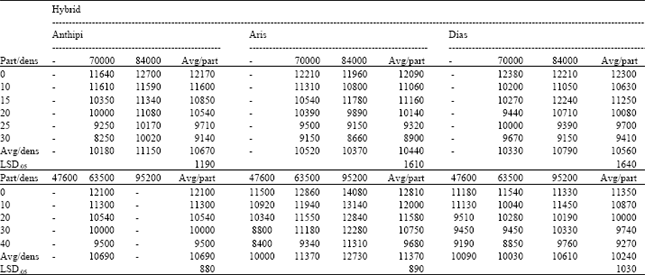 | |
In the area of Karditsa, the yield (y) of hybrid Anthipi was negatively correlated to the presence of inbred parent (x) and the equation of the regression line was: y= 12392-103.43x (R2=0.971, r= -0.985, at p<0.0001). For hybrid Aris the equation was: y= 12265-109.2x (R2=0.9475, r= -0.973, at p<0.001). For hybrid Dias the equation was: y= 12125-93.8x (R2=0.8847, r= -0.941, at p<0.005). In the area of Larissa, the yield of hybrid Anthipi was negatively correlated to the presence of inbred parent and the equation of the regression line was: y= 11988-65x (R2=0.9883, r= -0.994, at p<0.0001). For hybrid Aris the equation was: y= 12866-75.1x (R2=0.98, r= -0.99, at p<0.001). For hybrid Dias the respective equation was: y= 11304-52.9x (R2=0.9726, r= -0.986, at p<0.005). Pure hybrid seed partitions always yielded better than low purity seed partitions (for all hybrids in Table 2), since values were increased from low purity conditions to the pure hybrid. This impact can be attributed to low yielding ability of inbred line plants and to increased number of barren plants (barren plants and inbred line plants decreased mean of lower purity seed partitions). Barren plants increase with decreasing hybrid purity[28]. In this study, low purity conditions proved to be a major stress in comparison to the density stress[10].
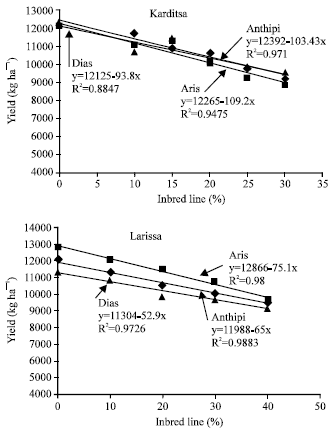 | |
| Fig. 1: | The negative correlation between yielding performance (kg ha-1) and the presence of inbred parent (%) for all hybrids, in the areas of Larissa and Karditsa, Greece |
The plant density influences mainly plant-to-plant variability, either in homogeneous F1s, or in heterogeneous maize genetic materials[10,14,27], with impact on yield performance even though considering autogamous species like tomato, where there are now available density-independent cultivars[29]. Increased plant-to-plant uniformity for yield per plant of maize hybrids, with decreasing the plant density was reported by Tollenaar and Wu[4], Vafias et al.[10] Similar impact of plant densities on plant-to-plant uniformity have been found for other traits, like dry weight and plant size[30] as well as when photosynthesis was involved[20]. The optimum plant density differed among hybrids and experimental areas, but in general, differences were not significant. For hybrid Anthipi, was found to be at 84000 plants ha-1 (in the area of Karditsa). For hybrid Dias was found between 63500 to 70000 plants ha-1 (combination of the two areas). Hybrid Aris was favored by high density in the area of Larissa (95200 plants ha-1) but in the area of Karditsa, the optimum was at 70000 plants ha-1.
Combination of data from two areas (Larissa and Karditsa) showed that hybrid Aris was an unstable genetic material in yielding performance, hybrid Anthipi was more stable and hybrid Dias was even more stable (Table 2). This was expected, because hybrid Aris is an older hybrid and in the opposite, hybrid Dias is a modern hybrid with greater adaptability. There was a tension showing that Aris could gain about 20% (statistically significant) in yielding performance at higher plant densities, Anthipi about 14% and Dias about 8%. The three hybrids had the same yielding performance in low plant density. Differences in grain yield between older and newer maize hybrids were shown to be a function of plant population density[11]. At lower plant densities, the differences between older and newer hybrids were very small, becoming greater as plant density increased[3,9,12]. The dependence of maize hybrid yield per unit area on plant density was also depicted by previously reported data[31,32]. In this study, very high optimum plant density (over 70000 plants ha-1) is being in agreement with that found by Tetio-Kagho and Gardner[31], Tokatlidis[14], Ipsilandis and Vafias[27]. Breeding in maize succeeded in developing hybrids with highly improved yield per unit area, through improvement in tolerance to stresses, including high plant densities[2,4,8,26]. Tollenaar and Wu[4] claimed that increased stress tolerance, combined with increased stand uniformity under stress conditions, would probably continue to provide the highest potential for yield improvement in maize. Troyer and Rosenbrook[32] suggested that selection under higher plant densities was a means to improve grain yield of maize, even though a negative relationship between competitive ability (performance under density stress) and yield in pure stands has been widely reported[33,34]. Conversely, for the same objective, Duvick[12] found it necessary to suggest selection under lower plant density environments (1 plant m-2) as a means of effecting future gains in yielding ability of maize hybrids. Without mentioning why and how to achieve such a goal, Duvick[12] probably had in mind the disadvantage for the farmers from the yield dependence on higher plant densities. Additionally, data presented by Tokatlidis[14] and Tokatlidis et al.[26] showed that recycling of a hybrid, aiming to improve its potential yield per plant via self-pollination and selection at the very low density of 0.74 plants m-2 on the basis of higher single plant yield and lower CV of individual plant yields, led to recycled hybrids which exhibited less density-dependence than their original hybrid.
The impact on leaf chlorophyll content: Leaf chlorophyll content (in SPAD units), was found to be better for pure hybrid entries in comparison to low purity seed partitions (Table 3). In the area of Karditsa, hybrid Anthipi reached an average of 51.2, hybrid Aris 57 and hybrid Dias 57.9 SPAD units. In the area of Larissa, the average values were 52, 57.3 and 57.5 SPAD units, respectively.
| Table 3: | Leaf chlorophyll content (in SPAD units) for the three pure hybrids (Anthipi, Aris, Dias), in two areas (Karditsa, Larissa), within the respective (6 or 5) seed partitions (part) and plant densities (dens), with average values (Avg), in comparison to their respective inbred parents |
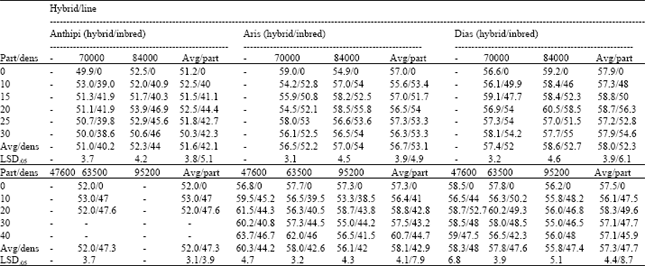 | |
This was in agreement with the yielding performance of pure hybrid. As hybrid plants and inbred line plants were measured separately, it was found that the reason for low leaf chlorophyll content measurements in plots of low purity seed partitions was due to inbred line performance. Inbred line performance could never overcome pure hybrid leaf chlorophyll content. In general, there were small (but statistically significant) differences between seed partitions (Table 3). The average plot leaf chlorophyll content (in SPAD units) decreased from 51.2 (pure hybrid) to 46.3 (highest level of inbred parent presence) for hybrid Anthipi in the area of Karditsa and from 52 to 47 in the area of Larissa. For hybrid Aris the respective values were decreased from 57 to 54.8 and from 53 to 52.7. For hybrid Dias the respective values were decreased from 57.9 to 56.2 and from 57.5 to 51.6. Plant density influence on leaf chlorophyll content was different in the area of Karditsa in comparison to the area of Larissa (Table 3). In Karditsa, leaf chlorophyll content was higher when plant density was increased. On the other hand, in Larissa, leaf chlorophyll content was higher when plant density was decreased. It is expected that, in low plant density, plants could express better their performance and be more vigorous[20,33] in a way that leaf chlorophyll content and photosynthesis must be higher in comparison to high plant density which proved to be a stress condition. Data from Larissa are in agreement with that statement. It is possible that in Karditsa, the two plant densities are too close (only 14000 plants ha-1) to distinguish such differences. Additionally 70000 and 84000 plants ha-1, are near the yielding performance optimum (as was found in this study and proposed by the company that owns these hybrids). The densities of 47600 and 95200 plants ha-1, are considered more extreme[33] and thus better for analyzing plant density effects. In general, seed purity effect was more significant than plant density stress.
The discussion on impure seed partitions presumes preliminary tests for distinguishing these partitions. In this study, the pure hybrid germinated better than low purity seed partitions. Seeds in 15°C germinated faster in comparison to 11°C, so that germination was completed within six days, for almost all hybrids. In 11°C, germination was completed after about 15 days. In this temperature, the pure hybrid was easily distinguished from low purity seed partitions in the sixth day of measurement for all hybrids, since there were significant differences.
Yielding performance of different genetic materials was influenced significantly by the seed purity conditions. Pure hybrid seed partitions yielded better than low purity seed partitions for all hybrids. There were differences in yielding performance between different plant densities, however, low purity conditions proved to be a major stress in comparison to the density stress. Extreme plant densities, away from proposed optimum (reference by seed producer) are better for analyzing plant density effects.
Leaf chlorophyll content (in SPAD units), was found to be better for pure hybrid entries in comparison to low purity seed partitions due to lower inbred line performance.
The practical conclusion of the findings demonstrate that seed purity is indeed a major stress in comparison to plant density and that an attentively conducted seed production program may insure yielding performance in the field for modern maize hybrids. Tokatlidis[14] and Tokatlidis et al.[26] consider that the development of density-independent hybrids is the solution for insuring high and stable grain yield in maize. In this study, density stress proved to be of minor importance in comparison to seed purity as was found also by Vafias et al.[10] As a final conclusion, we propose that attentively conducted seed production programs of maize hybrids that exhibit density tolerance (plus tolerance in various biotic and abiotic stresses) may insure high and stable yielding performance.
REFERENCES
- Fehr, W.R., 1987. Principles of Cultivar Development. Vol. 1, Macmillan Publishing Co., New York.
Direct Link - Tollenaar, M. and J. Wu, 1999. Yield improvement in temperate maize is attributable to greater stress tolerance. Crop Sci., 39: 1597-1604.
CrossRefDirect Link - Bonan, G.B., 1991. Density effects on the size of annual plant populations: An indication of neighbourhood competition. Ann. Bot., 68: 341-347.
Direct Link - Dwyer, L.M. and M. Tollenaar, 1989. Genetic improvement in photosynthetic response of hybrid maize cultivars, 1959 to 1988. Can. J. Plant Sci., 69: 81-91.
Direct Link - Nissanka, S.P., M.A. Dixon and M. Tollenaar, 1997. Canopy gas exchange response to moisture stress in old and new maize hybrids. Crop. Sci., 37: 172-181.
Direct Link - Duvick, D.N., 1984. Genetic Contributions to Yield Gains of U.S. Hybrid Maize, 1930 to 1980. In: Genetic Contributions to Yield Gains of Five Major Crop Plants, Fehr, W.R. (Ed.). ASA and CSSA, Madison, WI., pp: 15-47.
CrossRefDirect Link - Fasoula, V.A. and D.A. Fasoula, 1999. Honeycomb breeding: Principles and applications. Plant Breed. Rev., 18: 177-250.
Direct Link - Tokatlidis, I.S., 2002. The effect of improved potential yield per plant on crop yield potential and optimum plant population in maize hybrids. J. Agric. Sci., 137: 299-305.
Direct Link - Edmeades, G.O. and T.B. Daynard, 1979. The relationship between final yield and photosynthesis at flowering in individual maize plants. Can. J. Plant Sci., 59: 585-601.
Direct Link - Snedecor, G.W. and W.G. Cochran, 1980. Statistical Methods. 7th Edn., Iowa State University Press, Iowa, USA., ISBN-10: 0813815606, Pages: 507.
Direct Link - Tokatlidis, I.S., M. Koutsika-Sotiriou and A.C. Fasoulas, 2001. The development of density-independent hybrids in maize. Maydica, 46: 21-25.
Direct Link - Ipsilandis, C.G. and B.N. Vafias, 2005. Plant density effects on grain yield per plant in maize: Breeding implications. Asian J. Plant Sci., 4: 31-39.
CrossRefDirect Link - Troyer, A.F. and R.W. Rosenbrook, 1983. Utility of higher plant densities for corn performance testing. Crop Sci., 23: 863-867.
Direct Link - Tetio-Kagho, F. and F.P. Gardner, 1988. Response of maize to plant population density. II. Reproductive development, yield and yield adjustments. Agron. J., 80: 935-940.
CrossRef - Thomas, J.B., G.B. Schaalje and M.N. Grant, 1994. Height, competition and yield potential in winter wheat. Euphytica, 74: 9-17.
Direct Link