
A.M.R. Abdel-Mawgoud
Not Available
S.O. El-Abd
Not Available
A.F. Abou-Hadid
Not Available
Y.N. Sassine
Not Available
Asian Journal of Plant Sciences
Year: 2005 | Volume: 4 | Issue: 6 | Page No.: 619-624







ABSTRACT
Data from two greenhouse experiments on sweet pepper (Capsisum annuum.) cv. Mazurka were analyzed to investigate the effect of different shoot and root environments on the incidence of Blossom End Rot (BER) in the fruit. First experiment consisted of four combinations of treatments with two levels of EC namely high EC (HEC) 6.5 dS m-1 and low EC (LEC) 2.5 dS m-1 both under two levels of transpiration, high (HET) and suppressed (LET). Second experiment had four combination of treatments and consisted of two levels of volumetric water content in the root zone, high (HWC, 80%) and low (50%), with two fruit load treatments, normal (NFL) and manipulated (MFL). Analysis of data showed that high EC in the root zone had a strong effect on increasing incidence of BER under both transpiration rates. BER was positively correlated with weekly cumulative energy sum (average daily temperature x average daily radiation), initial fruit growth rate and average fruit size of healthy fruit. The data indicate that high average leaf area per growing fruit seems to be the central factor inducing BER. Therefore, it is suggested to prune old leaves of sweet pepper plants as being practiced with other greenhouse crops.
PDF Abstract XML References Citation
How to cite this article
A.M.R. Abdel-Mawgoud, S.O. El-Abd, A.F. Abou-Hadid and Y.N. Sassine, 2005. Blossom End Rot of Sweet Pepper Fruits in Relation to Different Shoot and Root Conditions. Asian Journal of Plant Sciences, 4: 619-624.
DOI: 10.3923/ajps.2005.619.624
URL: https://scialert.net/abstract/?doi=ajps.2005.619.624
DOI: 10.3923/ajps.2005.619.624
URL: https://scialert.net/abstract/?doi=ajps.2005.619.624
INTRODUCTION
Blossom End Rot (BER) is a physiological disorder caused by local deficiency in calcium[1] with symptoms being observed at the distal end of the fruit. In pepper as well as in tomato, the calcium concentration reduces from the proximal to the distal tissue of the fruit where the symptoms of BER occur[2]. Whilst supply and uptake of calcium are not limiting in most systems[3], the most significant factor here is its distribution once in the plant[4]. Investigation on the cause of BER in tomato show that low calcium supply or uptake as well as low transport of calcium to the fruit particularly to the distal fruit tissue, can induce BER even when the calcium status in the plant is high[5,6]. The Electrical Conductivity (EC) of the nutrient solution may also be important as it has been shown that increasing EC of the root zone not only reduces calcium uptake but also reduces xylem density within the fruit[3,7,8]. This leads to a subsequent increase in the incidence of BER. Another situation under which BER is common is when there are elevated rates of fruit expansion resulting from high rates of influx of photoassimilates. Thus it can be seen that any environmental condition that favors photosynthesis such as high temperature or high ambient light intensity, will lead to increase in incidence of BER[9].
Therefore, it is the purpose of this study to analyze the interaction between different shoot and root conditions on the incidence of BER in sweet pepper.
MATERIALS AND METHODS
In the two experiments, sweet pepper plants cv. Mazurka were grown in rockwool slabs in a one or two compartments of a multispan Venlo-type greenhouse in Wageningen, The Netherlands (52° N) from the second week of January to mid of August. The plants were grown in a density of 2.2 and 4.2 plants m-2 in the first and second experiment, respectively. First and last rows in the greenhouse were used as borders as well as the first and last plant in each row. Plants were trimmed weekly to two first order stems (twin branch system) with all side shoots stopped at one leaf and flower. The terminal flower was removed in all plants to support vegetative growth. In the first two weeks after planting, the plants were grown in 80% volumatric water content in the root environment followed by four weeks in 65%.
Greenhouse climate: Inside air temperature, RH% and CO2 concentration in the compartment as well as outside solar radiation were recorded every 5 min by a commercial Vitaco computer system (Hoogendoorn, ‘s Gravenzande, The Netherlands) and controlled by heating and ventilation. Day/night temperature set points were 22 and 19°C, respectively. No thermal screen was used.
Treatments: In the first experiment, two treatments of volumatric water content in the root environment in combinations with two fruit load treatments (total four treatments) were arranged in a Split Plot Design where water content was the main variable. Water content treatments (V:V) were High Water Content (HWC) 79-81% (control) and Low Water Content (LWC) 49-51% and started on the 9th of March. Water content and EC in the root zone were monitored by online two Grodan water-content sensors, one in each treatment, connected to the main greenhouse computer. Another Gordan hand water-content sensor was used to check other slabs once a week to ensure homogeneity in water treatment during the experiment. EC was monitored and kept in the range 3.0±0.5 dS m-1 by switching the irrigation from nutrient solution with EC 3.0 dS m-1 to nutrient solution with EC 1.0 dS m-1 whenever the EC in the root zone goes above 3.0 dS m-1.
Fruit load treatments were Normal Fruit Load (NFL) where all possible fruit set were allowed (control) and Manipulated Fruit Load (MFL) where only one flower per stem week-1 was allowed to develop to fruit (two fruits plant-1 week-1) and all other flowers and small fruits were removed. In MFL treatment, preference was given to the flowers on the main stem, however, if not available in anthesis stage, flower on the side shoot was chosen. Six and eight plants per plot were used for weekly fruit set measurements for NFL and MFL, respectively. Fruit set was recorded once a week until end of June and twice a week for normal fruit load to the end of the experiment.
Yield was monitored on 12 and 22 plants/replicate for NFL and MFL treatments, respectively.
Harvest was carried out twice a week when fruits were fully ripe (red color) and fruits were classified to marketable and non-marketable fruits.
Treatments were arranged in a Split Plot Design consisting of three blocks with water treatment is the main variable and fruit load treatment is the sub-main.
In the second experiment, two compartments were transplanted. In each compartment, with sweet peeper seedlings plants were fed from two tanks with two different levels of EC namely low EC (LEC) = 2 dS m-1 and high EC (HEC) = 6.5 dS m-1, each tank feeding half of the plants in each compartment. Both these treatments started four weeks after transplanting to ensure similar root development. At each irrigation cycle, the water drawn from the tank was adjusted automatically (if necessary) to the required EC level, by injection of concentrated solution (A/B tank system) and/or fresh water. The composition of the solution was adjusted twice monthly, following chemical analysis of some extracts from each EC treatment.
The greenhouse climate was controlled in order to manipulate potential transpiration without causing difference in photosynthesis rate. The first compartment was controlled according to the Dutch practice (high transpiration rate) and in the second compartment potential transpiration was limited not to exceed 0.15l plant-1 h-1. Transpiration in the first compartment was calculated according to the model described by Stanghellini[10]. Control of the transpiration rate was carried out by injecting vapor by means of high pressure misting system. In the greenhouse climate control program, the set point of windows opening for ventilation based on vapor pressure deficit was 25% smaller in the second greenhouse compared to the first one.
The experiment was designed in a Split Plot Design with three replicates, with transpiration as the main plot and EC as the sub-main plot. Statistical analysis was carried out using Genestat package.
RESULTS AND DISCUSSION
Blossom end rot tended to shorten the growth period of the fruit. BER fruits had an average growing period of 7 weeks compared to 8 weeks for normal fruits. The incidence of BER usually occurs in the stage of fast growing (1-2 weeks after fruit set)[11]. Therefore for an accurate investigation for any possible relation with BER incidence, number of BER fruits was correlated to any possible factor 5-6 weeks before harvesting these fruits.
Effect of energy sum: The incidence of BER in other crops such as tomato has been shown to be markedly affected by light and temperature independent of the calcium supply to the plant[9]. In this study the concept mentioned by Marcelis and Ho[12] of energy sum (average daily temperature X average daily solar radiation, °C.M J m-2 d-1) was used. However, a weekly cumulative of this energy is more representative for the effect on BER incidence. Indeed a clear trend to the incidence of BER and energy sum can be seen in Fig. 1, for all the treatments of the two experiments. A correlation between weekly cumulative energy sum and incidence of BER under normal EC level was found and can be expressed by the following formula:
| Y= 0.001X-0.06 (R2= 0.69) |
Where, Y is the number of fruits affected with BER (fraction of total yield) and X is the weekly cumulative energy sum (average temperature * Average solar radiation, °C.M J m-2 week-1).
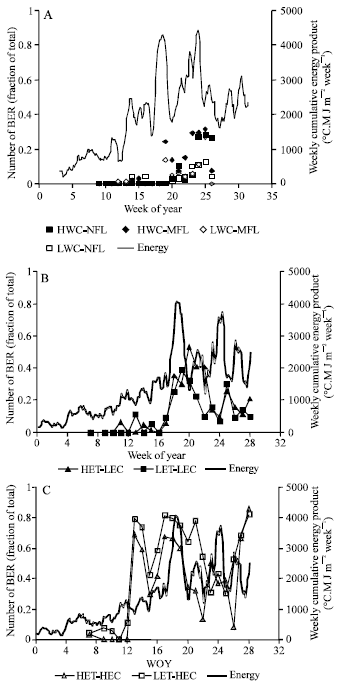 | |
| Fig. 1: | Weekly cumulative energy sum (°C.M J m-2 week-1) in the sixth week before harvest and number of BER fruits (fraction of total yield) in water content and fruit load treatments (A), Low EC (LEC) (B) and High EC (HEC), (C) under Low Transpiration (LET) and High Transpiration (HET) conditions |
According to this equation, it seems that there is a threshold of energy (612 °C.M J m-2 week-1) beyond it, incidence of BER occurs. The percentage of BER is also proportional to the peak of energy above the threshold. In case of HEC, the relationship with the energy product was not so clear like in case of LEC. It seemed that there was another factor interfering with the energy sum concept. It has been reported that increasing EC of the nutrient solution reduces calcium supply to the fruits and xylem density within the fruit[3,7,8] that leads to a subsequent increase in the incidence of BER and this can be applied in this case.
Effect of fruit load, initial fruit growth rate and average fruit fresh weight: Competition for assimilates and Ca is related to fruit load on the plant. It follows that BER incidence must have a relation with fruit load. Figure 2 shows that there is a certain level of fruit load on the plant beyond it, incidence of BER occurs.
It is clear from Table 1 that only NFL under LWC was significantly lower than the other treatments concerning calculated maximum growth rate (at inflection point). However, there was a tendency for a higher growth rate in MFL treatments and HWC treatments in general.
| Table 1: | Fitted parameters of Gompertz function to measured fresh weight of growing fruits (±Standard Error) and Maximum fruit growth (at the inflection point) |
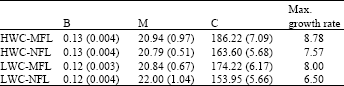 | |
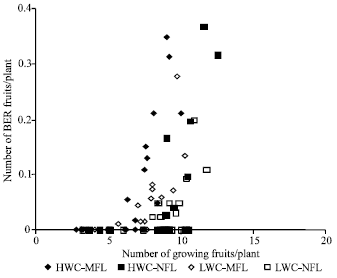 | |
| Fig. 2: | Number of BER fruits at harvest (fraction of total yield) and number of growing fruits on the plant six weeks before harvest in the four combinations of water content treatment in the root zone and fruit load treatments |
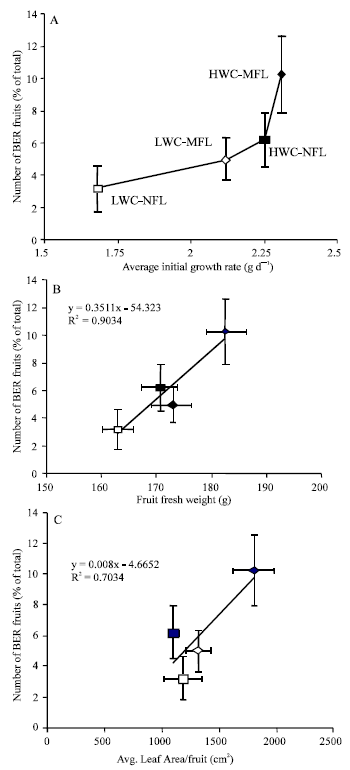 | |
| Fig. 3: | Percentage of number of BER fruits in relation to initial fruit growth rate during 0-17 days after fruit set (A), average fruit fresh weight of healthy fruits (B) and average leaf area per growing fruit (C). Symbols are the same for A, B and C. Error bars are twice standard error |
It was suggested previously that accelerated growth rate of the fruit may induce BER in tomato[13,14].
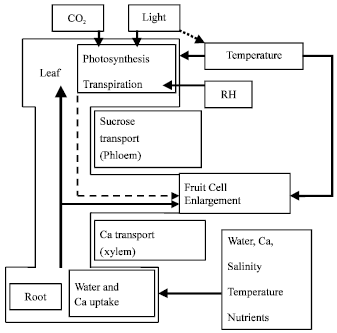 | |
| Fig. 4: | Schematic diagram explains the effect of different environmental factors on Ca transport in the plant and the incidence of Blossom End Rot (BER) (after Adams and Ho (1993)) |
Figure 3A confirms this suggestion where a clear positive relation between initial growth rate of the fruit (0-17 d after anthesis) and the incidence of BER exists.
Another positive relationship between average fruit fresh weight of healthy fruits and the percentage of BER incidence was observed (Fig. 3B).
Leaf area is an important factor in assimilate production (source) hence affecting the growth rate of fruits (sink). There was a positive relationship between the two parameters. The relationship indicates that when leaf area per growing fruit exceeds a certain level (~ 600 cm2) there is an increase of 1% in BER incidence for each 125 cm increase in leaf area per fruit (Fig. 3C).
BER in sweet pepper became visible from 3 weeks after anthesis. Over this period both the rate of fresh weight and dry weight gain in pepper fruit increased rapidly and reached the maximum rate[15]. This rapid initial growth rate can be increased further by increasing source-sink ratio (i.e. by fruit pruning). In tomato, the induction of BER coincided with a low Ca concentration during the period of rapid growth[7]. Similar positive relationship between initial fruit growth rate and incidence of BER was observed in this study. This also may contribute to the positive relationship between average fruit fresh weight of healthy fruits and the percentage of BER incidence. Similar relationship was also reported by Marcelis and Ho[12] with different sweet pepper cultivars and fruit treatments. They also reported a negative relationship between fruit size and calcium concentration in the fruit. This explains the positive relationship between fruit size and incidence of BER that was observed here. Similar to observation in sweet pepper[12] and in tomato[9] the incidence of BER showed a positive correlation with the product of temperature and radiation. The latter two factors act in two different directions that induce BER (Fig. 4). The two factors enhance assimilate production (faster fruit growth rate) and also transpiration from the leaves (more Ca going to the leaves), hence bigger chance of BER incidence. Reducing transpiration by high humidities favors the build up of root pressure[5] a process that force the xylem stream being forced into those areas of tissue not usually well supplied. As calcium is mobile in this stream it follows that this process is important in calcium nutrition to the fruit. However, in contrary to SCHON[16], who successfully used anti-transpirant, De kreij and Huys[17] couldn’t diminish the percentage of sweet pepper fruits affected with BER by regulating plant transpiration using high humidity. The results reported here were in agreement with the latter since there was no effect of LET treatment in diminishing the incidence of BER. However, in this presented experiment of high EC and climate manipulation (LET), the higher number of BER fruits seems to be mainly due to the higher number of fruits on the plant[2] which increased the competition among the fruits for calcium supply. This was clear in the smaller cumulative number and weight of BER fruits in HET-HEC treatment where the total number of fruits was smaller hence the competition must have been less. Another factor that can affect this high percentage of BER under HEC treatments is the high ratio of potassium and magnesium to calcium in the root zone. When the concentration of potassium and magnesium in the root zone are high this has an antagonistic effect on calcium uptake, which may increase the likelihood of BER occurring[18].
It can be concluded that some desired agricultural practices that increase fruit quality such as fruit pruning might also induce the incidence of BER. It is very important to keep a reasonable leaf area per fruit when working with fruit pruning since leaf area is the central parameter for both photosynthesis and transpiration. High EC in the root zone induces the incidence of BER regardless of climate conditions.
REFERENCES
- Bangerth, F., 1979. Calcium-related physiological disorders of plants. Ann. Rev. Phytopathol., 17: 97-122.
CrossRefDirect Link
Aroshini Reply
I am a 4th year student in a srilankan campus and I am doing a research in bell pepper field. So I want datasets for calcium deficiency in bell pepper fruit. Can I have your datasets if they are still available?
Editor
Thank you for your interest in the research on the Blossom End Rot of Sweet Pepper Fruits. We encourage you to conduct your own research in the bell pepper field to investigate calcium deficiency in bell pepper fruit. We believe that your study could contribute valuable insights into this area of research.
We wish you all the best in your research endeavors.