
Asfaw Adugna
Not Available
Asian Journal of Plant Sciences
Year: 2005 | Volume: 4 | Issue: 3 | Page No.: 316-322







ABSTRACT
In this study the two pollination control systems, i.e. Cytoplasmic Male Sterility (CMS) and the use of Chemical Hybridizing Agents (CHA), were compared to discover their efficiency for the sterility reaction and their impacts in hybrid seed production. The result has shown that both systems were equally effective in their sterility performance. However, CHA resulted in reduced out crossing percentage due to female sterility, germination percentage and hundred seed weight of the hybrid seed due to toxicity. Overall the CMS system was better than the CHA system (based on the particular CHA used in this study) for attributes related to hybrid seed production.
PDF Abstract XML References Citation
How to cite this article
Asfaw Adugna, 2005. Behavior of Bread Wheat (Triticum aestivum L.) Male Sterilized with Cytoplasmic-genetic and Chemical Systems and Their Impact in Hybrid Seed Production. Asian Journal of Plant Sciences, 4: 316-322.
DOI: 10.3923/ajps.2005.316.322
URL: https://scialert.net/abstract/?doi=ajps.2005.316.322
DOI: 10.3923/ajps.2005.316.322
URL: https://scialert.net/abstract/?doi=ajps.2005.316.322
INTRODUCTION
Seed production is a vital part of any hybrid programme with percent seed set on a male sterile plant an important economic factor in seed production[1,2]. For successful adoption of hybrid variety technology, the essential prerequisites are the availability of proper pollination control system to enable production of commercial quantities of hybrid seed and existence of adequate degree of heterosis to make the hybrid commercially viable[3,4]. Hybrid wheat requires a transformation in flower behavior from self to cross-fertilization[5]. Hybrid seed can be produced using either a Cytoplasmic-Genetic Male Sterility (CGMS) restorer system or by using Chemical Hybridizing Agents (CHAs)[6,2]. A requirement of any method of obtaining male sterility in wheat is the control of pollen production in all florets including those produced by early or late tillers[7]. Hybrid seed production in wheat using CMS is a three lines system and involves three steps[8]: Multiplication of CMS (A-lines); Multiplication of maintainer (B) and restorer R-lines and production of hybrid seed (A x R). In using the chemical system to produce hybrids, two lines would be necessary, a P-line (pollinator) for use as the male parent and C-line (chemical male sterile) used as the female parent[9]. The effective use and versatility of CHAs will depend on genotype-chemical, environment-chemical and genotype-environment-chemical interactions[8].
Kihara[10] was the first to report male sterility in wheat from the original intergeneric crosses he made between Aegilops caudata and Triticum aestivum in 1935 where wheat acted as a female parent. However, the first workable CMS, which is still used for commercial production, was derived from T. timopheevi cytoplasm[11].
Most CMS sources confer complete male sterility on T. aestivum nucleus but they are associated with some defects. Not only are many cytoplasms inducing male sterility in wheat: a range of other effects has also been known to CMS lines and the hybrids produced from them[12]. Pickett[7] classified these effects as deleterious, neutral and beneficial. According to Ghiasi and Lucken[13] most of the Triticum (other than T. timopheevi) and Aegilops cytoplasms have deleterious effects on various traits. Therefore, a majority of these cytoplasms are not usable, due to associated biological penalties in terms of reduced vigor, abnormal plant morphology, zygote elimination and delayed maturity[14].
The observation of complete pollen sterility on plants treated with Maleic Hydrazide by Chopra et al.[15] opened the first door towards the possible use of CHAs for hybrid seed production in wheat. Later, several companies like Rohm and Haas, Shell and Monsanto have carried out extensive work on development of gametocides[16]. The requirements of an ideal CHA according to Chopra et al.[15], Stoskopf[17] and Singh[18] are causes only male sterility without affecting female fertility; non-mutagenic; consistently produce complete male sterility; be easy and economical to apply; have minimum side effects to humans. From the practical view point, CHAs must be effective if applied over a range of crop growth stages, on different genotypes and in different environments; should effectively sterilize both early- and late-formed flowers on tillers, the chemical should be in a form that can be safely sprayed on the designated female strips without danger of drift to the adjacent male rows; should have a minimal impact on such factors as spike development and emergence from the boot, leaf chlorosis and heading date or productivity.
Gametocides with an absolute effect are the golden dream of every breeder[19]. Unfortunately, no CHA used until recently appears to have achieved all the requirements for effective CHA[14]. Although many chemicals have been investigated for use as male sterilants, each of them was associated with at least some shortcomings. Therefore, like the CMS system CHAs have also been blamed for one or more of the following side effects: Lack of flexibility in the time of spraying (Ethrel)[20], shortening of the treated plants (Ethrel, RH531)[21-23], delayed flowering of the CHA treated male sterile lines (Ethrel), inhibition of ear emergence (Ethrel)[21], incomplete male sterility (Ethrel), Less effective for early tillers (RH532)[22], Genotype specificity, narrow application window, large dose rates and seasonal differences in sterilizing efficiency (DPX3778)[20,24,25], sterilization of female gametophytes (Maleic hydrazide, RH531, RH532, SD84811, WL84811)[2,22,26,27], phytotoxicity, toxicity to humans and animals and toxic residues in the F1 seed, etc. A further demerit of the use of CHAs is that it would add to the cost of F1 hybrid wheat seed[28]. Therefore, this experiment was carried out to discover the behavior of bread wheat male sterilized with T. timopheevi based CMS system and CH9832 based CHA system for hybrid seed production and their impacts on the resulting hybrid seed.
MATERIALS AND METHODS
The research was carried out at the experimental farm of the Department of Plant Breeding, Punjab Agricultural University, Ludhiana, during the winter seasons of 2000-2001 and 2001-2002 under irrigated condition.
Plant material: The plant material consisted of 20 male sterile lines (A-lines) (carrying T. timopheevi cytoplasm, undergone at least four backcrosses); 20 maintainer (B-lines) corresponding to the 20 male sterile A-lines used and 20 restorer (R) lines (Table 1 and 2). These materials were used in various combinations for hybrid seed production.
| Table 1: | CMS lines (carrying T.timopheevi cytoplasm) used for hybrid seed production |
 | |
| Table 2: | List of R-lines used for hybrid seed production |
 | |
Lay out of seed production plots: Two types of hybrid seed production plots were laid out:
| Set 1: | This set of plots consisted of a pair of CMS lines with a row of restorer genotype on either side |
| Set 2: | This set of plots consisted of a pair of corresponding B-lines (non-sterile) with a row of restorer genotype on either side. The same cross combinations were maintained in the two sets, the only difference being that set 2 contained male fertile versions (isolines) of CMS lines used in set 1. The lines in set 2 were later CHA sterilized for hybrid seed production. To ensure synchronization of flowering each combination was grown in three temporal staggering (Fig. 1). |
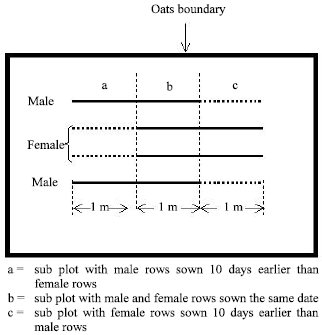 | |
| Fig. 1: | A schematic representation of the lay out of individual seed production plots |
Each plot consisted of 4 rows (2 outer males and 2 inner females) each of 3 m length. Each 1 m length corresponded to the above-mentioned temporal staggering. The 4 rowed 1 m plot length (sub plot), which showed adequate synchronization, was used for recording observations related to seed production attributes.
Besides hybrid seed production plots, control genotypes comprising 20 B-lines, not treated with CHA were included in the trial. Each plot had been planted with plant-to-plant spacing of 4 cm and row-to-row spacing of 23 cm. The different hybrid seed production and control plots were sown in a randomized complete block design with 3 replications.
Each plot was isolated from other plots with thick sown rows of a tall variety of oats (Avena sativa) to avoid pollen contamination from the neighboring plots. Natural wind pollination was supplemented with hand shaking. Management practices were as per the recommendation of the area.
CHA spray: The B-lines in set-2 were sprayed with CH9832 at a dose of 1000 ppm when ear primordial length in the primary tillers was from 7 to 10 mm (corresponding stage 22 on Zadok et al.[29] growth scale). CH9832 was obtained from the directorate of wheat research, Karnal, India. Tween 20 was used as a surfactant. These plots represent the CHA based seed production plots.
Observations, efficiency indices and data analysis: Observations on days to 50% anthesis and plant height were recorded on female and male lines of CMS/CHA sterilized plots as well as controls.
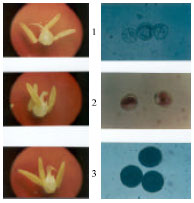 | |
| Fig. 2: | Plates 1, 2 and 3 show the microscopic view of the pollen grains and the male and female floral organs on either side for the samples taken from the CHA-sterilized, CMS and control plots, respectively |
Moreover, observations on sterility rate, out crossing rate, germination percentage, thousand grain weight and percent of effective seeds related to F1 seed production attributes and were recorded on CMS and CHA treated females only.
For the purpose of calculating sterility percentage five ears were bagged in each CMS and CHA sterilized plot just before flowering. The mature, bagged spikes from each plot were collected and threshed together. Sterility percentage was calculated by deducting the selfing percentage from hundred. Where, selfing percentage is equal to the mean number of seeds per bagged spikes divided by the mean number of seeds per spike on the control multiplied by 100. In this, the seeds per spike averaged from five random B-line spikes served as control. Moreover, for calculating the out crossing percentage, 10 open pollinated ears from the female line were sampled from each CMS or CHA sterilized plot and the 10 ears were threshed together. Seeds counted and from the total number of seeds the mean number of seeds per open pollinated spike was worked out. Out-crossing percent was worked out as (the mean number of seeds per open pollinated spike minus the number of seeds per bagged spike) divided by the number of seeds per spike on the control and multiplied by 100. In other words it is percentage of florets setting seed in hybrid seed production plots after deducting for possibility of selfed seeds. The selfed seed data was the same as was used for working out the sterility percentage in the previous attribute. Percent Effective Seeds was calculated as crossed seed production (Out-crossing percentage) multiplied by the probability of germination.
Pollen viability was observed by staining with Acetocarmine. Figure 2 shows photographs of pollen grains from CMS lines, CHA treated lines and normal wheat lines and the effect of CMS and CHA systems of male sterility on floral organs.
Finally, the data were subjected to analysis of variance for factorial experiment in RBD.
RESULTS AND DISCUSSION
Analysis of variance: In these hybrid combinations, the CMS lines PBW396, HD2652 and DL802-3 did not give adequate levels of sterility. These hybrid combinations and their CHA based combinations had to be excluded from subsequent observations and analysis. The sterility systems and genotypes represented the two known causes of variation in this experiment. The analysis of variance (Table 4 and 5) showed sterility systems (CMS vs. CHA) to be a highly significant source of variation for days to anthesis, plant height, out crossing percentage, thousand- grain weight of crossed seed and effective crossed seeds. Genotypic differences were significant for days to anthesis, plant height, out crossing percentage and thousand grains weight of crossed seeds. The relative magnitude of mean squares showed sterility systems to have a much greater effect on various parameters as compared to genotypes. The interaction (Sterility system x Genotypes) was significant for days to anthesis and plant height indicating differential response of genotypes to sterility systems.
Effect of male sterility systems on days to anthesis: In case of days to anthesis (Table 3) CMS lines were earlier by a little more than one day. However, there was no significant difference on this character for the CHA sterilized lines and the control. The days taken by CHA sterilized lines was at par with control. The interaction mean square was significant indicating differential response of genotypes with respect days to anthesis caused by CMS or CHA.
Effect of male sterility systems on plant height: The systems mean showed that CHA-treated lines were significantly shorter than the control whereas the CMS lines were at par with control (Table 3). Several researchers[9,20,21,30] reported similar results.
| Table 3: | Effect of sterility systems on various attributes of the female lines and the control |
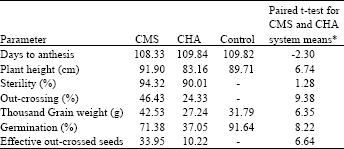 | |
| *Mean values for each character are found in Adugna et al.[31] | |
The short height in CHA treated lines was most probably due to the toxic effect of the CHA used because yellowing of plants in response to CHA treatment was observed during the course of the experiment. According to Rowell and Miller[9], lowered height of female lines in case of CHA induced sterility may possibly be a morphological advantage to hybrid seed production as that could increase the level of out crossing.
Level of male sterility in CMS vs. CHA treated lines: Both systems were equally effective for male sterilizing efficiency (Table 3). The mean sterility percentages were 94.32 and 90.01% for CMS and CHA, respectively. Many genotypes showed 100% sterility by Adugna et al.[31] . Figure 2 shows a similar result.
Out crossing potential of CMS and CHA facilitated male sterility systems: In this experiment, sterility systems had drastic effect on out crossing percentage (Table 3). On the average, genetic system of male sterility showed an out crossing percentage almost two times higher than the CHA system (46.43% in CMS and 24.33% in CHA). Genotypic differences were also significant among the females within each group in the ability to set seed from air-borne pollen which was in agreement with the results of Bitzer and Patterson[32]. Genotypic specificity of toxic response to CHA is also indicated. The mean out crossing percent of CMS system was 46.43 and 24.33 for the CHA system.
| Table 4: | Mean squares from the analysis of variance for T. timopheevi based CMS lines and lines treated with the chemical hybridizing agent CH9832 |
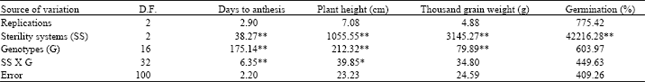 | |
| *, ** Significant at 0.05 and 0.01 levels of probability, respectively | |
| Table 5: | Mean squares from the analysis of variance for T. timopheevi based CMS lines and lines treated with the chemical hybridizing agent CH9832 |
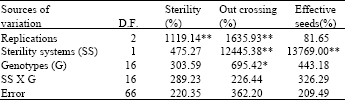 | |
| *, ** Significant at 0.05 and 0.01 levels of probability, respectively | |
Although the causes of poor out crossing are considered to be influenced by the morphology and biophysics of pollen and the effect of climate[33], the reduced seed set on the CHA treated plants in the present study is most likely due to female sterility. Some differences in synchronization of flowering, in spite of the three staggered sowing provided may also have contributed to variation in seed set. Efforts were made to utilize hybrid seed data only from synchronized plots.
Effect of male sterility systems on thousand grain weight: The seeds produced from CMS lines were bolder but to some extent dented than the control whereas CHA treated lines produced shriveled and smaller seeds than the control as a result of toxicity of the CHA. Systems factor was highly significant (Table 4). Genotypic mean squares were highly significant showing availability of heritable differences among genotypes for this character. The mean thousand grain weight of the CMS system (42.53 g) was greater than the control (31.79 g) but the CHA system was lower than the control (Table 3).
Effect of male sterility systems on germination percentage: Systems factor was highly significant for germination percent (Table 4). Both CMS and CHA based crossed seed showed lower germination than the control. On the average, the germination of the seed from plants treated with CHA was almost half that of the germination level of CMS seeds. The overall systems mean showed the highest germination percentage value (91.64) in controls followed by the CMS based crosses (71.38) (Table 3). CHA based crosses showed the least germination percentage (37.05) as the seeds were shriveled. Lower germination in crossed wheat seed was also reported by Johnson and Lucken[34]. Shriveling in turn seemed to be due to toxicity in case of CHA treated seeds. However, it was hard to explain the cause of denting in case of CMS seeds, which were otherwise the boldest of the three groups. Several authors have given similar reports. For instance, Rai[35] had reported frequent occurrence of kernel shriveling and pre-harvest sprouting in the F1 seeds produced by the T. timopheevi system. He concluded that these deleterious effects were caused by the interaction of the T. timopheevi cytoplasm and the wheat nuclei.
Effect of male sterility systems on percent effective seeds: Effective seeds index based on out crossing percentage and germination ability was used to assess the overall hybrid seed production potential of the two systems. The grand mean percentage of effective out crossed seeds from CMS lines (33.95%) was three times more than that from CHA treated lines (10.22%) (Table 3).
CH9832 represented the best available CHA, which had been specifically standardized for use on wheat after tested in the North Western Penesular Zone (NWPZ) of India for 2 years. None of the internationally used CHAs was accessible for comparative studies. However, the T. timopheevi based lines were well advanced (Table 5).
The absence of such experimental comparisons in wheat imparts a wider relevance to this study. The study has clearly shown the CMS system to be superior for hybrid seed production attributes.
Although wheat hybrid production has been attempted in the past, it was for the most part abandoned due to the difficulties inherent to its development[36]. Among the disadvantages usually given regarding economical hybrid wheat seed production are the high seeding rate requirements and the problems of seed production arising from the cross pollination potential[11]. The main difficulty is the low seed set on the male sterile parent in seed production fields that could result in prohibitively costly hybrid seed[7,37]. As the success of hybrid wheat depends greatly on the female seed production[38], cross-pollination of sufficient magnitude and reliability is essential to the economic production of hybrid seed. However, usually this is not met because of various factors. Due to genotypic differences with respect to those factors seed set can range from less than 10% to more than 70% on male sterile lines[37]. Therefore, improvement of cross-pollination potential is likely considering the variability present in existing wheats[39].
ACKNOWLEDGMENTS
The author acknowledges Dr. N.S. Bains, Asst. Wheat Breeder, Punjab Agricultural University, Ludhiana, India for his unreserved help during the course of the research. The financial assistance by the Agricultural Research and Training Project (ARTP) of the Ethiopian Agricultural Research Organization (EARO) in the form of scholarship is duly acknowledged.
REFERENCES
- Zadoks, J.C., T.T. Chang and C.F. Konzak, 1974. A decimal code for the growth stages of cereals. Weed Res., 14: 415-421.
CrossRefDirect Link