
Wafaa Kamal Taia
Not Available
Asian Journal of Plant Sciences
Year: 2005 | Volume: 4 | Issue: 2 | Page No.: 184-206







ABSTRACT
Not Available
PDF Abstract XML References Citation
How to cite this article
Wafaa Kamal Taia, 2005. Modern Trends in Plant Taxonomy. Asian Journal of Plant Sciences, 4: 184-206.
DOI: 10.3923/ajps.2005.184.206
URL: https://scialert.net/abstract/?doi=ajps.2005.184.206
DOI: 10.3923/ajps.2005.184.206
URL: https://scialert.net/abstract/?doi=ajps.2005.184.206
INTRODUCTION
Taxonomy is the science of grouping organisms into different categories according to their physical characters. This grouping, or classification, should be based on homology i.e. the individuals gathered in the same group must shared characteristics that have been inherited from a common ancestor. Thus, to study the modern trends in taxonomy we must go through the history of classification and how this branch has developed with the development of peoples, instruments and techniques. We must keep in our minds that taxonomy today is a reflection of the past, meanwhile the systems of classification reflect both needs, level of knowledge, philosophical concepts and available technology of each historical period.
HISTORY OF CLASSIFICATION
Greene[1], Copeland[2], Croizat[3], Lawrence[4], El-Gamal[5], Brush[6], Sokoloff[7] and Search in internet in History of Plant Taxonomy (up to date).
Classification of organisms have been started from the beginning of human existence basing on their need, shelter, food and medicine. Whenever it started, plant taxonomy has at least six distinct periods:
1- Preliterature, 2- Ancient Literature, 3- Medieval or dark ages 4- Renaissance, 5- Theory of Evolution, 6-Taxonomy Revolt. Each of these periods has its own characteristic features and achievements. So, if we go quickly through each of these periods we can recognize how plant taxonomy progress and develop.
Preliterature period: In this period people were very close to the earth, got their food from hunting and gathering. They knew many plants that are used for food and medicine. They were practical plant taxonomists, they described the plants, classified them according to wether they are useful or harmful, identified them and put them in a named category so it could be easily referred to.
Ancient literature: This is the period of the ancient greeks in which their conclusions reached by reasoning instead of analysis of observations. They noted the differences between external and internal organs. They classified plants by form into trees, shrubs, undershrubs. They, also, recognized annuals, biennials, perennials and floral morphology. Meanwhile, they described many medicinal plants which considered the basic source of information for more than 1500 years.
Medieval (dark ages): In this period a little has been done in plant taxonomy except that of Albertus Magnus who recognized monocots and dicots, vascular and nonvascular plants.
Renaissance: This period had influenced by the following: 1- Printing Press was invented, 2- Individuals had confidence to attempt original work, 3- Navigation enabled collection of plants from all over the world. For that, this period was an active period of learning and exploration, meanwhile many large volumes about plants and their uses (herbals) were produced. It was the initial efforts of the ancients to structure and order flowering plant diversity. As well as, many natural and well defined genera and families were established beside naming of plants as proposed by Carl Linnaeus (father of plant taxonomy). The beginning of numerical taxonomy (phenetics) has been started by Adanson. An early thinking of evolution has been achieved by Lamark as well as the use of internal structures beside the external ones in classification has been done by de Candolle.
Theory of evolution: Prior to the theory of evolution by Charles Darwin, a geologist called Charles Lyell proposed a theory of geological gradualism says that “there is a slow, continuous changes produced features we see today”. Thus, Darwin thought that the earth was much older than 6000 years and that evolutionary change must occurred through gradual accumulated differences. He provided conclusive evidence that evolution of life forms has occurred and proposed natural selection as the mechanism responsible for these changes. In the same time Alfred Wallace developed a very similar evolutionary theory and both of them were presented at the meetings of the London Linnaean Society. From that time taxonomists started to investigate which characters are primitive and which considered advanced. They tried to represent syntheses of data from all areas of botanical science such as anatomy, genetics, physiology, paleobotany, chemistry and palynology. Thus taxonomy enter a new phase which is phylogenetic classification which reflect distances and relationships between organisms. Thinking has been turned to the creatures of our time must had common ancestors in the past and accordingly, phylogenetic trees have been postulated.
Taxonomy revolt: From the preceding survey of the development of plant taxonomy, we can observe that the rules of classification are purely human invention and arbitrary, simply they are matter of choice. Thus, taxonomists started to revolute with the revolution in the microscopes and modern techniques. The invention of the scanning microscopes made things easy to know and internal structures become more obvious. DNA sequences and the study of comparative genome organization in plants made it more precise in detecting the relationships between the taxa. Simply, we can say that new era of taxonomy has been started.
MODERN TRENDS IN PLANT TAXONOMY
Within the last fifty years, plant taxonomists have begun to look for more precise techniques in order to understand the relation between the genera and the families. A lot of questions have been brought in mind, which is more advanced and which are less? What is the origin of these plants? Which more related and which must kept apart? And many other questions are kept in mind. These have made the taxonomists to go farther inside the internal structures of the plants and especially the chromosome structure, DNA sequences and genome structure. This what we called molecular taxonomy. Data obtained from this study are compared with those obtained from other tools of taxonomy and all these data have used for cladistic analysis and phylogenetic interpretation. In this review, we have to go through the most recent works in plant taxonomy and systematic.
Morphology: Plant morphology in all its aspects; from
micromorphology to embryology, palynology, seed, seedling, fruit, floral, stem and leaf anatomy, epicuticular secretions; is still hugely important. With proper weighting, morphological characteristics remain, till now, the most valuable tools in the phylogenetic tree. Here we have to mention some recent treatments and monographs as examples of the use of plant morphology and anatomy in recent classifications.
Vegetative morphology: Vegetative morphology must be the first step in plant identification. So the neglect of vegetative characters is one of the most serious errors in the history of classification and has done much to delay the achievement of a natural system. White[8] reported the reason of neglecting the vegetative characters by many taxonomists to the modular construction of the vegetative parts of higher plants i.e. the plant body is made up of indefinite number of repeating units without definite numbers of parts. Kellogg[9] suggested that taxonomists must return back to morphological taxonomy. Since the early 1980`s, plant systematics has focused on molecular phylogenetics, which had led some scientists to bemoan the demise of morphological taxonomy and the loss of morphological expertise. Recent works in using vegetative morphology in the subscription of the families of flowering plants are those of Bell[10], Brummitt[11], Watson[12], Takhtajan[13], Thorne[14] and Taia[15,16].
According to Taia[15] the leaves of the Papilionoideae have been divided into seven categories according to their trichome types, while Taia[16] found five types of leaf morphological characters, according to them tribe Trifolieae has been reclassified (Table 1).
Floral morphology and anatomy: The most active works in this field are those of Dickison[17-20] who studied the floral anatomy of the families Caryocaraceae (1990), Aristolochiaceae (1992) and Styraceae (1993) beside the floral anatomy of the species Sanango racemosum of the Gesneraceae (1994). Those of Endress,[21-27], on primitive angiosperms (Magnoliidae) (1990, 1994) tropical flowers evolution and diversity (1994), the evolution in Ranunculaceae (1995) and those of family Dilleniaceae (1997).
Endress has proposed an evolutionary prospects for the next century in a chapter of the book entiteled “Evolution and diversification” (1997). Erbar and Leins[28-32] were mainly on the floral anatomy of Sympetalous plants.
Link[34-36]. has studied the nectarines in family Geraniacea, while Ronse and Smets[37-46] have studied the androecium morphology and ontogeny in many families and genera especially those of the Magnoliaceae and Ranunculaceae. Bayer[47,48] used the inflorescence characters in the systematics of Malvales; Balthazar and Endress[49] studied the floral bract function in two genera of Chloranthaceae); Deroin[50,51] studied the vascular architecture of flowers in families Annonaceae and Magnoliaceae.
From the most interesting recent work in this line is that of Bruckner[52], who used the number of carpels as a useful character in classifying the Papaverales. He used gynoecial features like shape of the stigma, zones of dehiscence, structural of the placental regions, vascular pattern, ontogeny and tetratological transformation in comparing between members of Capparaceae, Papaveraceae and Fumariaceae.
| Table 1: | Macromorphological characters on leaflets of the studied taxa within tribe Trifolieae[33] |
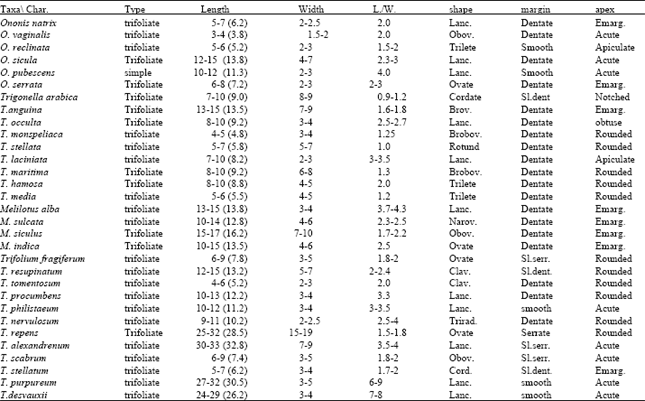 | |
| The measurements by mm.of at least ten middle leaflets, minimum-maximum (mean) Abbreviations used: Lanc.=lanceolate Obov.=obovate Brov.=broadly ovate Narov.=narrowly ovate Clav.=clavate Trirad.=triradiatus; Sl.dent.=slightly dentate Sl.serr.=slightly serrate; Emarg.=emarginated | |
Taia and Sheha[53] found that members of the Fumariaceae have different stigma than those of the Papaveraceae that can be an easy way to distinguish between their taxa.
All of these, beside many others, used the structures of the flowers and the ontogeny of each floral whorl as a tool for understanding the relation between the studied taxa.
Palynology: Now a days, palynological investigations become a popular tool in plant taxonomy, especially with the invention of the high resolution power microscopes as the scanning and transmission electron microscopes. The basic of this branch is Erdtman`s[54] Handbook of Palynology and its second edition by Nilson and Praglowski[55]. The most informative recent works in this field are those by Abu-Asab and Cantino[56,57], on members of Lamiaceae; Abu-Asab et al.[58], on the genus Caryopteris of the Labiatae; Carrion et al.[59], on the genus Coris of the Primulaceae; Ferguson et al.[60], on family Leguminosae; Nowicke[61-63], on the order Caryophyllales and included families beside the Crotonoideae of the Euphorbiaceae.
Other taxa in which pollen studies clarified the phylogenetic position of the group are those of the genus Phyrma and its relalives[64]; Globularieae and Selagineae[65]; members of the families Apocynaceae and Periplocaceae[66]; members of the Ranunculanae[67]; the Paleobotanical work on the early Angiosperm pollen from the Cretaceous[68,69]; genus Triplostegia of the Valerianaceae[70]; members of family Calyceraceae[71]; family Goodeniaceae[72]; the pollen morphology of the genus Oryza (Poaceae)[73] and genera of family Caryophyllaceae and tribe Trifolieae[74,75].
Within the Caryophyllaceae four pollen types have been recognized which can be used in the taxonomy of the family. Taia and Sheha[53,76,77] studied the pollen characters in members of the Boraginaceae and they recognized three pollen groups which can help in understanding the phylogeny of the family. They studied, also the pollen grains of Atriplex species and they described it by being eupalynous. While Taia[75,78] had studied the pollen morphology in species of Amaranthaceae and those species of tribe Trifolieae.
| Table 2: | Palynological characters as shown by both light and Scanning electron microscopes within Members of Tribe Trifolieae[75] |
 | |
She found three pollen types and five subtypes within members of tribe Trifolieae which can help in understanding the evolutionary line in the group (Table 2).
These works used the pollen characteristics like, the structure and ornamentation of the wall, polarity, symmetry, shape, size and apertures in a phylogenetic order of the studied group.
Embryology: In the last twenty years, numerous investigations of the embryology of dicotyledons have been most helpful in the proper placement of various taxa. The use of the number of integuments, seed coat and nucellus type in the phylogeny of the dicotyledons has been proposed by many authors. Image of double fertilization by Friedman[79-83] and he continued his search with his colleague Williams[84], as it first evolved in a common ancestor to the gnetophytes and the angiosperms. The second fertilization product was diploid and yielded a supernumerary embryo.
Subsequent evolution led to the establishment of an embryo-nourishing, nuclear endosperm and eight-nucleate embryo sac. The origin of the endosperm in flowering plants have been discussed, repeatedly by Friedman[79,83,85], Friedman and Williams[84], Grosniklaus and Schneitz[86], Bowe et al.[87], Chaw et al.[88], Baroux et al.[89], Soltis et al.[90] and Williams and Friedman[91].
Donald and Todd[92] had gone through the history of biology and how Darwin`s works and his elucidation of the alternation of generations had influenced Mendel`s genetic studies.
The International Conferrence on Plant Embryology[93] had covered the researches done on plant reproduction from Mendel to molecular biology.
Seed and fruit morphology: Barthlott[94] has given an account of seeds and small fruits morphology as important tool in plant taxonomy, especially after the invention of the transmission microscopes which opened new vistas in this study. Recent investigations in this study are those of Chuang and Ornduff[95] on the seeds of Menyanthaceae; Setten and Kock-Noorman (1992) on fruits and seeds of Annonaceae; Rohwer[96-98] on fruits and seeds of Oleaceae; Ryding[99] on pericarp structure of Pogostemoideae of family Lamiaceae; Spjut[100] on the systematic treatment of fruit types.
Taia[74], Taia and Mohammed[101,102] have studied the seed surface of the Caryophyllaceae and they found great variation in seed ornamentation between the the three subfamilies, meanwhile the seed characters revealed that members of subfamily Paronychioideae is homogenous and their classification into two groups, as previously suggested, is not supported.
Ahmed and Taia[103] found that pod character in the genus Medicago can help in the subscription of the group.
| Table 3: | Stigma and leaf characters in the studied taxa[53] |
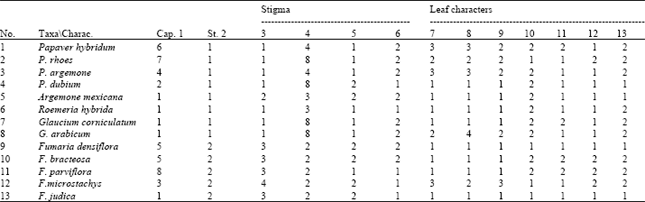 | |
| Characters studied: 1-Capsule shape:1-linear 2-obovate 3-obcordate 4-clup-shape 5-obtuse 6-rounded 7-globular 8-rugose 2-style: 1-abscent 2-present 3- stigma shape: 1= orbicular lobed disc 2=crater-shape 3=lever-like 4=flat 4-number of stigma 5-presence of hairs in the scapsule: 1=present 2=abscent 6-presence of stomata in the capsule: 1=present 2=abscent 7-hair density on the leaves: 1=abscent 2=sparse 3=hairy 8-hair type: 1=abscent 2=simple-pointed 3=ramified 4=glandular 9-hair wall: 1=abscent 2=smooth 3=granulated 10-type of stomata: 1=diacytic 2=anomocytic 11-shape of epidermal cells : 1=isodiametric 2=elongated 12-shape of anticlinal walls: 1=straight 2=wavy 13-epicuticular secretions: 1=present 2=abscent | |
Bruckner[104] studied the seed structure in family Ranunculaceae; Cameron and Chase[105] on the seed morphology of Vanilloid orchids; Mirle and Burnham[106] on asymmetrically winged samaras from the Western Hemisphere; Taia[107] on the seed morphological features in tribe Trifoleae.
Karyomorphology: As now defined, karyomorphology involves more than just the number of chromosomes in a given plant; it also involves their size, morphology and internal anatomy. This taxonomical tool considered as one of the most important way in the circumscription of many taxa. Greilhuber[108] pointed to the necessity of using karyomorpholgy and chromosome banding in taxonomy. From the recent works in using karyomorphological data in taxonomic studies are those Snow and Goldblatt[109] in species of the genus Ticodendron; Benko-Iseppon and Morawetz[110,111] on the karyosystematics of the genera Sambucus and Viburnum (Adoxaceae) and the cytological features of order Viburnales; Graham et al.[112] on the chromosome numbers in the genera Sonneratia and Duabanga (Lythraceae); Baum and Oginuma[113] on members of family Bombacaceae; Turner[114] on those of the Caryophyllales; Okada[115] on family Chloranthaceae; Heubl and Witsuba[116] on the cytology of Nepenthes and Stace et al.[117] on the cytoevolution in family Epacridaceae. After 1997, molecular investigations of DNA and RNA sequences were the subjects of research.
Anatomy: The internal structures of the plants have added a lot in understanding the relationships between the taxa. The organization of the vascular system in stem and leaf, petiole and nodal anatomy, leaf vasculature and architecture and epidermal studies are considered important characters in phylogeny and taxonomy. Barthlott[118] used the epicuticular wax secretions in the systematics of Caryophyllales. He continued using the epicuticular secretions in the classification of the Rununculiflorae[119] and furthermore in the classification of angiosperm[120].
Dickinson[120] studied the stem anatomy of the genus Aphanopetalum and found that it can help in the taxonomy of the genus. Meanwhile, Al-Shammary and Gornall[121], used the trichome anatomy in the systematic of Saxifragaceae; Gibson[122] used the internal structures of the stem in the delimitation of the genera in family Cactaceae. Vestures in the Myrtales, Gentianales and Fabales have been studied by Jansen et al.[123].
While the works done on the internal structures of the leaves were less than those on stems. However, some articles on trichomes, stomata, cuticles and leaf architexture have been useful in helping to establish relationships. Among those are the works done by Baranova[124,125], on the epidermis of Austrobaileya (a) and the comparative stromatographic studies of angiosperms; Wilkinson[126,127], on the leaf anatomy of both Pittosporaceae and Pterostemonaceae; Kohler[128], on the leaf venation in Buxaceae and Simmondsiaceae; Al-Shammary and Gornall[121] on the trichomes in the Saxifragaceae; Jorgensen[129], in stomatal myrosin cells in Cariaceae; Klucking[130,131], on the leaf venation patterns in Euphorbiaceae; Christopher et al.[132] on cuticular features in Lauraceae; Baas[133], on the Boraginaceae; Noshiro and Baas[134], on the internal structures of both the leaves and stems of the genera of family Cornaceae and its allies.
Ye[135] studied the vascular tissue differentiation and pattern formation in plants; Al-Turkey et al.[136] used the primary vasculature in the leaves of family Chenopodiaceae and its applications for systematics and evolution and Taia and Sheha[53] and Taia[107] on the microcharacters of the leaves in tribe Trifoleae (Table 3 and 4).
| Table 4: | Micromorphological characters on leaflets of the studied taxa within tribe Trifolieae[33] |
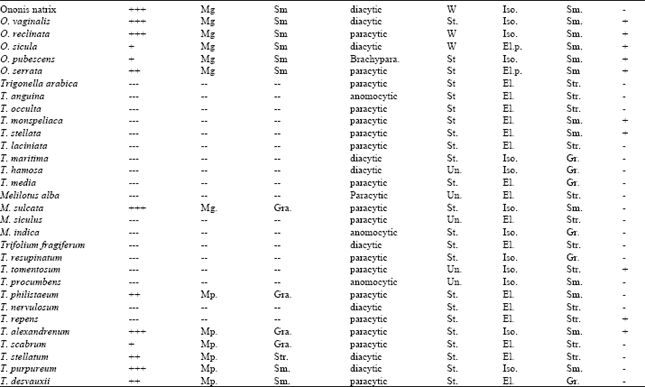 | |
| Characters: 1= hair density 2= hair type 3= hair wall 4= stomatal type 5=anticlinal wall 6=shape 7=cuticle 8=wax secretions - = absent + = present ++ = dense +++ = woolly. Abbreviations used : Mg.=multicellular glandular, Ug.=unicellular glandular, Mp.=multicellular pointed; Sm.=smooth, Gra.=granulate, Str.=striate; Brachypara.=brachyparacytic; W=wavy, St.=straight, Un.=undulate; Iso.=isobilateral, El.p.=elongated pentagonal, El.=elongated; Gr.=granular | |
Wood anatomy gained more intersest in this field than the other structures for phylogenetic purposes. The characters which are of taxonomical importance in this field are: cell types, vessel elements, vessel length and width, types of perforation plates and lateral wall pitting and thickness. From the most important works in this field are those of Carlquist[137-159] on the wood anatomy of families Lamiaceae, sympetalous families, Cucurbitaceae, Chloranthaceae, the genus Eupomatia, Aristolochiaceae, Sabiaceae, Caryophyllaceae, Rananculaceae, Glaucidiaceae, Berberidaceae, Menispermaceae, Akaniaceae and Bretschneideraceae, the genus Pentaphragma, Buddlejaceae, Resedaceae, Portulaccaceae and Hectorellaceae, Caricaceae and the genera Petiveria and Rivina of Caryophyllales, Agdestis, Schisandreaceae, Stegnosperma and Basellaceae.
All of these, beside many others have added a lot to taxonomy. They found that wood structure and type can help in the taxonomy and the phylogeny of the studied taxa. Baas et al.[160] have studied the wood anatomy of the Dicotyledones and its application in angiosperm classification. Baas and Wheeler[161] pointed to the importance of wood anatomy in biodiversity.
Chemotaxonomy and serology: Comparative biochemists have continued to present useful information for phylogenists. The chemical compounds and presents in some taxa can help in understanding their relations with another taxa. For example, the phytochemical study of the family Podostemaceae[162] showed that there is no relationship with the genus Hydrostachys. Gottlieb et al.[163] found that the primitive members only of Hamamelididae, Dilleniidae and Rosoideae have galloyl esters in their tissues. Chemistry and micromolecular studies in the Fabaceae by Gottlieb et al.[164] indicated a probable derivation from Sapindales.
Works concerning phytochemical survay can be showed by Bohm and Chan[165] on the flavenoids in the genus Greyia; the distribution of iridoids and other compounds in Loganiaceae and other families of Asteridae by Jensen[166]; the chemosystematic overview of Magnoliidae, Ranunculidae, Caryophyllidae and Hamamelididae by Gottlieb et al.[167]; the chemotaxonomy of the genus Samango of Gesneriaceae by Jensen[168]; the secondary compounds of Ranunculanae by Jensen[169]; the chemotaxonomy of three closely related genera of the Lamiales by Denton and Smith[170]; the phytochemistry and chemotaxonomy of the Boraginaceae by Hegnauer[171]; the study of phytochrome B and E in early angiosperms by Mathews[172]. Jensen[173,174] studied the chemotaxonomy of the genus Nuxia (Buddlejaceae) and the chemical relationships of Polypremum procumbens, Tetrachondra hamiltonii and Peltanthera floribunda. Von Poser et al.[175] have studied the distribution of iridoids among the tribes of Bignoniaceae and the data obtained can help in the taxonomy of the family. Albach et al.[176] studied both the embryological and biochemical contents in the evolution of the asterids. Jensen and Schripsema[177] studied the chemotaxonomy and pharmacology of Gentianaceae.
Jensen et al.[178] studied the chemotaxonomy of the Oleaceae and found that the biosynthesis and distribution of iridoid can be used as taxonomic markers. Ronsted et al.[179] used the chemical constituents in the genus Plantago as basis for its taxonomy and evolution. Albach et al.[180] used the iridoid glucosides of Paederota lutea to understand the relationships between Paederota and Veronica.
Serological investigations are less common in taxonomical works, from these are Jensen et al.[181] on which they found that all taxa of high legumin similarity belomg to Malvanae, Rutanae and Violanae, with Euphorbiaceae closely allied with Utricales, Flacourtiaceae and Passifloraceae but even more similar to Thymelaeaceae. Jensen[182] also found that legumin is the main seed-storage protein in Ranunculaceae, supporting the classification based on other molecular, morphological and chemical data.
Pfister et al.[183] has developed analytical and diagnostic methods for Larkspur (Delphinium spp.) alkaloids, including development of immunogenic alkaloid-protein conjugates and antibodies for a systematic approach to the taxonomic classification of the genus.
Paleobotany: Paleobotanical works are those concerned with fossils. Their discoveries add much to phylogeny and geography of the flowering plants. From these studies we can predect the direction of evolution and plant phylogeny and origin. Two important books dealing with the origin and diversification of flowering plants are those edited by Taylor and Hickey[184] and by Iwatsuki and Raven[185]. Cornet and Habib[186] described angiosperm- like pollen from the upper Jurassic of France and speculated that the angiosperms separated from ancestors of Bennetitales and Gnetales before the Carnian age of the Triassic or the Permian.
Crane et al.[187] reported on the origin and major diversification of the angiosperms in the early Cretaceous. Kranz et al.[188] tried to find the origin of land plants from the study of the ribosomal RNA gene sequences among charophytes, bryophytes and vascular plants. Graham[189] found that the green algae is an evolutionary transition to land plants.
Van Boskirk[190] studied the paleofloristics of the lower Campanian Eagle Formation of Wyoming and Montana and he listed fossils of members of flowering plants beside those of conifers and ferns.
From the most interesting complication on the origin and development of grasses is that written by Jacobs et al.[191]. As they proposed that it is originated in Paleocene in S.America. It was rare until opening of forests in early to mid Tertiary with simultaneous increase of C3 grasses. Origin of C4 grasses was in middle Miocene and they spread over C3 grasses by late Miocene.
Graham[192] Studied the Neotropical paleobotany, an Oligo-Miocene palynoflora; while Graham et al.[193] continued this study from the Middle Eocene Saramaguacan Formation of Cuba.
From the more abundant representation of extant dicotyledonous families are known from the Tertiary by Manchester[194]. Schonenberger and Friis[195] studied the fossil flowers of ericalean affinity from the Late Cretaceous of Southern Sweden. Klavins et al.[196] studied the anatomy of Umkomasia (Corystospermales) from the Triassic of Antarctica.
Jordan and Macphail[197] found a Middle-Late Eocene inflorescence of Caryophyllaceae from Tasmania, Australia.
Phylogeny: Phylogenetic works are those concerning with the data obtained by paleobotanists to discover the relations between plants. Works done in plant phylogeny are numerous from the time of Darwin, but we have to mention the most important, recent ones in this field. Among the most informative longer publications pertinent to the better understanding of dicotyledon phylogeny are: Carnivorous plants: Phylogeny and evolution, by Albert et al.[198]; Vascular Plant Families and Genera, by Brummitt[11]; Advances in Labiatae Science, by Harley and Renolds[199]; Advances in Legume Systematics Part 4 The Fossil Record by Herendeen and Dilcher[200]; Erdtman`s Handbook of Palynology, edited by Nilson and Praglowski[55]; Seeds of Woody Plants of North America by Young and Young[201]; Wood Anatomy of the Rosaceae by Zhang[202]; The Genera of the Umbelliferae: A nomenclature by Pimenov and Leonov[203]; The Cruciferae of Continental North America by Rollins[204]; Asteraceae: Cladistics and Classification by Bremer[205]; Caryophyllales: Evolution and Systematics by Behnke and Mabry[118]; Systematics and Evolution of Rununculiflorae, by Jensen and Kadereit[169]; The phylogeny of the genus Rubus of the Rosaceae by Alice and Campbell[206]; The Anther: Form, Function and Phylogeny by D`Arcy and Keating[207]; Flowering Plant Origin, Evolution and Phylogeny, by Taylor and Hickey[184]; Evolution and Diversification of Land Plants by Iwatsuki and Ravin[185]; Diversity and Classification of Flowering Plants by Takhtajan[13]; The phylogeny of the Rubioideae by Anderson and Rova[208]; Seeds: Ecology, Biogeography and Evolution of Dormancy and Germination by Baskin and Baskin[209]; The phylogeny of subfamily Ixoroideae (Rubiacea) by Andreasen et al.[210]; Seed Plant Phylogeny inferred from all three Plant Genomes: Monophyly of extant gymnosperms and origin of Gnetales from conifers, by Chaw et al.[88]; The Classification and Geography of the Monocotyledon Subclasses Alismatidae, Liliidae and Commelinidae by Thorne[211]; Evolutionary Origin of the Endosperm in Flowering Plants by Baroux et al.[89]; the evolution of genome size in the angiosperms by Soltis et al.[212]; the evolution of the sexual process in ancient flowering plant lineages by Friedman and Williams[84] and on fossil evidence for supporting current molecular phylogeny by Wang[213].
Molecular biology: The biggest change in plant classifications happened since the highly informative results produced by the molecular taxonomists through analyses of DNA sequence data from the chloroplast gene rbcL. Also, the data obtained from chloroplast restriction analysis of the inverted repeat and nuclear rRNA sequencing. These data together with morphological and other data from the other tools of taxonomy have made great change in the world of taxonomy.
Before reviewing the most recent works in this field, Rieseberg and Soltis[214], Doyle[215] pointed that the gene trees are mostly based on single gene whether of chloroplast or nucleus chromosomes and that most molecular trees based on sequence data, no matter how parsimonious, are single-character trees. This means that the phylogenetic trees derived from molecular data only will be taken with precaution. Only phylogenetic trees derived from molecular data which are congruent with macromolecular ones, we can accept such approach as a powerful new tool for phylogeny.
Among the most instructive molecular articles on the dicotyledon orders and higher categories are: Donoghue et al.[216], on Dipsacales; Giannasi et al.[217], on Caryophyllales; Olmstead et al.[218,219], on Asteraceae; Rettig et al.[220], on Caryophyllales; Price and Palmer[221], on Geraniales; Chase et al.[222], on Magnoliideae; Manos et al.[223], on Hamamelididae; Rodman et al.[224-226], on Capparales; Manhart and Rettig[227], on Caryophyllales; Savolainen[228,229], on Celastrales; Nickrent and Soltis[230], on angiosperms; Olmstead and Reeves[231] and Olmstead et al.[232], on Scrophulariaceae; Bayer[233], on Antennaria; Clement and Mabry[234], on Caryophyllales; Conti et al.[235,236], on Myrtales; Gadek et al.[237], on Sapindales; Gustafson et al.[72], on Asterales; Hibschjetter and Soltis[238], on Saxifragales; Nickrent[230], on Santalales and Rafflesiales; Plunket et al.[239,240], on Araliales; Sytsma et al.[241], on Utricales; Sytsma et al.[241,242], on Myrtales; Vincent et al.[243] eudicots; Ablett et al.[244], on Hernandia albiflora; Manos and Steele[245], on higher Hamamelididae; Soltis et al.[246,247], Albach et al.[248], on Asteridae; Al-verson et al.[249,250], on Malvales; Bayer et al.[251], on Malvales; Bayer and Starr[252], on Asteraceae; Xiang et al.[253], on Cornales; Les et al.[254], on Nymphaeales; Magallon et al.[255], on eudicots; Bakker et al.[256], on Pelargonium; Chaw et al.[88], on seed plant; Bazzaz[257], on plant biology in the future; Federici et al.[258] on Oryza officinalis (monocotelydon); Hsu et al.[259], on tandem families (monocotyledon); Jorgensen et al.[260], on Oxytropus campestris.
Diversity and adaptation: By the last twenty years, a great deal of concern throughout the world regarding the loss of biological diversity[261]. They return this loss to the change in the ecosystem. The diversity found within species is the ultimate source of biodiversity at higher levels. Genetic variation, population dynamics and genetic population structure all together shape and influence the way a species interacts with its environment and with other species. The term adaptation means the ability of the organism to overcome environmental changes. To measure the adaptation, there is a term used called Key Adaptation (KA) or Key Innovation (KI) refers to attempts over the past 50 years to relate patterns of evolutionary change and diversification to the appearance of particular traits.
For that scientists have turned to study the ways of respond of the species to the environmental conditions, from these works are: Evans[262], on Brassica campestris; Coleman et al.[263], in plants; Hogan et al.[264], on tropical forest; Cheplick[265], on Amphibromus scabrivalvus; Pigliucci et al.[266], on Arabidopsis; Williams et al.[267], on Pennisetum setaceum on Hawaii; Mazer and Gorchov[268], on distinguishing between genetic and environmental causes on progeny phenotype; Nicotra et al.[269], on tropical Piper; Cordell et al.[270], on Hawaiian tree species; Ashton[325], on species in West and East Malaysia; Soltis et al.[90], on floral diversity; Allen[271], on plants.
| Table 5: | Characters studied and their status in the three habitats[16] |
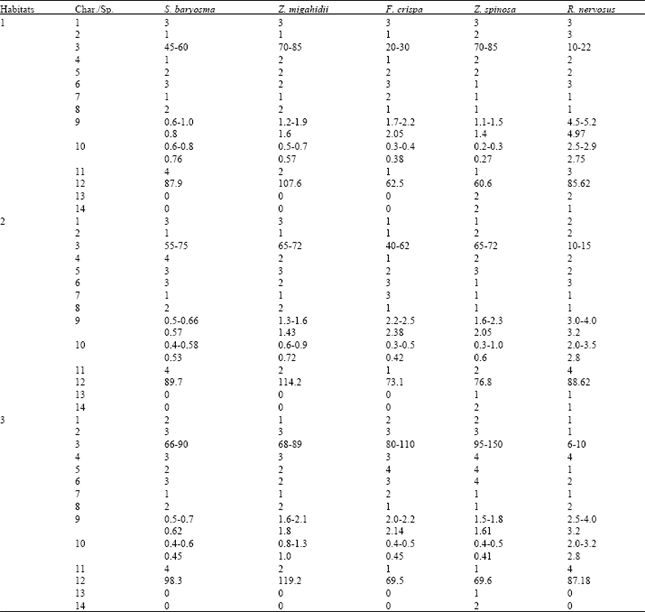 | |
| Characters: 1- Density 1= low, 2=moderate, 3=dence; 2- Appearance 1=dry, 2=slightly flourished, 3=flourished; 3-Length of the shoot system in cm; 4-Colour of the shoot system 1=whitish green, 2=green, 3=bright green 4=olive green; 5- Density of branches 1=unbranched, 2=branched, 3=densely branched, 4=very densely branched; 6-Leaf colour 1=pale green, 2=green, 3=olive green, 4=dark green; 7-Leaf margin 1=entire, 2=undulated, 3=sinuated; 8- Leaf apex 1=acute, 2=rounded; 9- Leaf length in cm.; 10- Leaf width in cm.; 11- Leaf shape 1=lanceolate, 2=ovate-lanceolate, 3=obovate, 4=broadly-obovate; 12-Plant moisture content (percentage); 13=Fruit 0=absent, 1=present, 3=dense; 14- Fruit shape 0=absent, 1=flat, 2=globular | |
Taia and El-Olayan[272], studied the effect of habitats on the phenotypic characters of three wild plants grown in El-Riyadh city in Saudi Arabia. Their results revealed that habitats have insignificant effect on the leaf characters. The main effects were on fruiting stage, length of the shoot system, colour of the plants and water content.
Taia and El-Ghanem[16], continued the study on the effect of habitats on both the mineral contents and the phenotypic characters of five widly distributed species in El-Riyadh city in Saudi plants. They found that, the leaf characters can be used as an effective taxonomic ones, while the mineral contents are greatly affected by both habitats and seasonal variations.
Ecotaxonomy: The ability of the plant to produce different phenotypes under different environmental condition which we called phenotypic plasticity has been an object of evolutionary and ecological studies since long time ago. Pigliucci[266] said that In recent years,a new dialog between organismal biologists and researchers interested in uncovering the mechanistic details of physiological and phenotypic responses has yielded several new insight.
These responses must affect the taxonomical characters of the plants (Table 5). Moreover they will affect the genome structure of them. Hoffman et al.[273] pointed to the selection on allozymes as well as quantitative traits may only occur under specific conditions and the responses to toxins illustrate how adaptive changes can be based on major genes with polygenic modifiers.
Stewart and Nilsen[274] studied the phenotypic plasticity within two isolated vaccinium macrocarpon, as they highly respond to edaphic factors, by usin RAPD (random amplifier polymorphic DNA) profiling. They found that the two isolates have different different plastic responses and one of them has higher molecular diversity. Delavega[275] returned the shaping of the population and species gene pools in response to environmental challenges to genetic adaptation. Jasienski et al.[276] have founded that there is genetic basis for phenotypic plasticity. More recent works have been done in this field (see diversity and adaptation).
Thus, we have to consider how much the environment may affects speciation and may cause to the presence of new species.
Cladistic analysis: Increasing use has taken place of computers for data storage and analysis during the past twenty years. Data derived from all tools of taxonomical investigations has to be analysed mathematically and cladistic trees have to be drawn. Burger[277] has written a strong critique of the Hennigian cladistics that so dominates modern-day taxonomy. Despite of the critisism of using cladistic analysis in taxonomy, cladistic methods have become a most useful technical tool for clarifying intrafamilial relationships.
From the excellent articles on the non-homology of vascular organization in monocotyledons and dicotyledons is Tomlinson[278], who listed the numerous characteristics that readily distinguish the two groups from each other. Brummitt[279,280], Brummitt and Sosef[281] and Sosef[282] have written tellingly on the excesss of the cladistics in regards to paraphylesis. Paraphyletic groups are the inevitable result of evolution. Thorne[211] wrote his own view in that taxa should be monophyletic but that the size of the phyletic gap between families should be the major consideration in their possible recognition.
Some of the more informative cladistic studies are: Anderberg[283], on Ericales Anderberg and Stahl[284], on Primulales; Hufford[285], on Rosidae; Hufford and Dickison[286], on Cunoniaceae; Lammers[287], on Campanulaceae; Karis et al.[288], on Cichoroideae; Karis[289,290], on Asteroideae; Graham et al.[112], on Lythraceae; Hershkovitz[291], on Portulacaceae; Hill and Jordan[292], on Nothofagus; Hoch et al.[293], on Onagraceae; Judd and Kron[294], Judd et al.[295], on Ericaceae according to morphological characters; Judd et al.[296], a preliminary phylogenetic analyses to angiosperm family pairs; Judd and Manchester[297], on Malvaceae according to different taxonomical tools; Kadereit et al.[298,299], on Papaveraceae; Rodman[300], on Caryophyllales; Chappill[301], on Leguminosae; Endress and Albert[302], on Apocynaceae; Gustafson and Brewer[303], on Asterales; Loconte et al.[304], on Ranunculaceae; Struwe et al.[305], on Gentianales; Albert and Stevenson[306], on Nepenthales; Smith[307], on Gesneriaceae; Struwe and Albert[308,309], on Gentianaceae; Zavada and Kim[310], on Ulmaceae; Hufford[311], on Hydrageaceae; Kron[312], on Rhododendroideae; Schlaurer[313], on carnivorous families; Kim and Jansen[314], on Oleaceae; Simmons and Hedin[315], on Celastraceae; Benton[316], on the stratigraphic data on the history of life; Chaw et al.[88], on seed plant phylogeny; Williams and Friedman[91], on angiosperm.
The obtained dendrogram separate the genus Ononis, as proposed by other taxonomic tools while the rest of the genera have been subdivided into two other subtribes.
Host-parasite relationships: Thorne[14] has pointed to the necessity of using the host-plant relationship and their predators or parasites in phylogeny. Ackery[317,318] studied the hostplants of the nymphalid butterflies and the hostplant utilization by African and Australian butterflies. Swensen[319] has found suggestions of multiple origins for actinorhizal symbiosis, although some groupings of actinorhizal taxa seem significant, including Cowantia, Purshia, Cercocarpus and Dryas of the Rosaceae; Rhamnaceae and Elaeagnaceae of the Rhamnales and Alnus of the Betulaceae with the Casuarinaceae and Myricaceae. De Souza et al.[320] found that the response of a host plant to gall-inducing insects varies both among and within plants. Nickrent[321] provided general synopsis of the current state of understanding of parasitic plant phylogeny by using modern analysis techniques. He concluded that parasitism has arisen independently ten times in each parasitic family i.e. each family is monophylatic. He found DNA sequences and molecular data answered long-standing questions in parasitic plant phylogeny.
TAXONOMY YESTERDAY
TODAY AND TOMORROW
Historically, Plant taxonomy start so simple as the needs of peoples to those plants. Gradually, this branch begin to be in the form of scientific works, as plant identity, distribution, morphology and physiology were emphasized. With the invention of the microscope, scientists were able to see small processes: internal structure, development and the stages in cell division became important subjects for plant study. Historical paleobotany concerned the evaluation of the progression of vegetation in a given location as the environment changed. It dealt with the slow changes in vegetation in a given region, the evolution of various taxa through geological time and the evolution of life. Thus, the fields of plant taxonomy were active. Evolution was approached by assuming the plants were adapted to their environment. With the discoveries of basic principles of inheritance and evolution by natural selection, the science of genetics and evolutionary biology flourished. There was much fascination with the development of common morphological traits in similar environments across phylogenetically disparate taxa. Thus, the investigations of paleobotanists provided a firm foundation for plant biology.
In the second half of the 20th century Molecular biology revolutionized the study of plants at all levels by discovering the structure of DNA and RNA, the steps in protein synthesis. Taxonomists, evolutionists, ecologists, physiologists and developmental biologists are now using molecular techniques and are discovering many responses and mechanisms that were not accessible in the past. It is now possible to identify, with much precision, the particular genes responsible for traits. As well as, with molecular techniques, scientists can introduce or eliminate genes for specific traits. Using these advanced techniques we may also alter the present taxonomy and phylogeny and, as the differences and similarities among taxa are modified by human action, we can create new species.
The environment in the future may be quite different from that of today. All indications suggest that the environment likely to be more variable than at present. According to many models, temperature and CO2 levels will rise. Furthermore, it is assumed that night temperature will increase disproportionally[322]. Although the interaction between the genotype and environment is complicated today, it will be more complicated in the future depending on the changing environments that the plant will face. Bazzaz and Stinson[323] stated we must remember that the environment has a direct influence on the genotype, in turning on and off genes. Also, we do not know which subset of genotype or environment will impact the most strategies of variation and fitness. All expectations suggest that the role of environmental variation may be increased. Bazzaz[257] has suggested that, according to the environmental changes, broad-niched species apparently respond well to global changes will be impacted less than narrow-niched species. Thus, it is expected that the increase in the environmental changes will to eliminate narrow-niched species. And species which are genetically variable and/or plastic in their response to environmental change will be favored[324].
From these previous works, we can say that taxonomy is a branch of science concerning with establishing all types of relationship among living organism. Also, it concerns with utilizing these relationships as the basis for systems of classification. Plant taxonomy, with its related studies of plant exploration, floristics, phylogenetics and phytogeography, is a vibrant and rapidly advancing discipline and one that is great practical relevance in the race against time to document, understand, conserve and utilize the world`s diminishing stock of botanical diversity.
So, we can expect that taxonomists have to go through all the informative tools, we must not forgot morphological taxonomy. We have to learn molecular techniques and try to discover the mechanisms of development and the control of developmental processes. This does not means that the basic structure of plant taxonomy will change, just we have to explore phylogenetic structures and developments through the study of the genome structures. Molecular study, alone, will not give a precise idea about evolution. Evolution is the cause of a natural system of classification; natural classification will represent the hierarchy of evolution. Classifications are based on features that seems to reflect common ancestry.
Ecology will become more important in taxonomy than before as the change in environments throught the coming years will lead to the extension of some species and appearance of new ones. We have to study plant diversity and the way of both internal and external adaptation according to environmental changes. Broad-niched species will dominate, while narrow-niched ones will eliminated. New species will arise according to both environmental changes and gene modifications by molecular techniques. Thus, we have to go further in investigating the change in our flora, how much our plants can cooperate with environmental disorders and to state new more stable taxonomic characters. Gene bank for the narrow-niched species must be done. We have to gather all the data from all the aspects of taxonomy, beside those derive from molecular study and ecological changes in order to understand the relationships between the taxa.
REFERENCES
- Taia, W.K. and W.M. El-Ghanem, 2004. Effect of habitats on both the phenotypic characters and mineral contents of five wild species in El-Riyadh city. Pak. J. Biol. Sci., 7: 1399-1403.
CrossRefDirect Link - Taia, W.K., 2004. Leaf characters within tribe Trifolieae (Family Leguminosae). Pak. J. Biol. Sci., 7: 1463-1472.
CrossRefDirect Link - Taia, W.K., 2004. Palynological study within tribe Trifolieae (Leguminosae). Pak. J. Biol. Sci., 7: 1303-1315.
CrossRefDirect Link - Al-Shammary, K.I.A. and R.J. Gornall, 1994. Trichome anatomy of the Saxifragaceae S.I. from the Southern hemisphere. Bota. J. Linn. Soc., 114: 99-131.
Direct Link - Al-Turki, T.A., K. Swarupanandan and P.G. Wilson, 2003. Primary vasculature in Chenopodiaceae: A re-interpretation and implications for systematics and evolution. Bot. J. Linn. Soc., 143: 337-374.
CrossRefDirect Link - Ye, Z.H., 2002. Vascular tissue differentiation and pattern formation in plants. Ann. Rev. Plant Biol., 53: 183-202.
CrossRefDirect Link - Jensen, S.R., H. Franzyk and E. Wallander, 2002. Chemotaxonomy of the Oleaceae Iridoids as taxonomic markers. Phytochemistry, 60: 213-231.
Direct Link - Price, R.A. and J.D. Palmer, 1993. Phylogenetic relationships of the geraniaceae and geraniales from rbcL sequence comparisons. Ann. Mssouri Bot. Garden, 80: 661-671.
CrossRefDirect Link - Chase, M.W., D.E. Soltis, R.G. Olmstead, D. Morgan and D.H. Les et al., 1993. Phylogenetics of seed plants: An analysis of nucleotide sequences from the plastid gene rbcL. Ann. Mo. Bot. Gard., 80: 528-580.
CrossRefDirect Link - Rodman, J.E., K.E. Karol, R.A. Price and K.J. Sytsma, 1996. Molecules, morphology and Dahlgren's expanded order capparales. Syst. Bot., 21: 289-307.
CrossRefDirect Link - Gadek, P.A., E.S. Fernando, C.J. Quinn, S.B. Hoot, T. Terrazas, M.C. Sheahan and M.W. Chase, 1996. Sapindales: Molecular delimitation and infraordinal groups. Am. J. Bot., 83: 802-811.
Direct Link - Janet Jorgensen, L., S. Ivana, B. Christian and C. Elena, 2003. Implications of ITS sequences and RAPD markers for the taxonomy and biogeography of the Oxytropis campestris and O. arctica (Fabaceae) complexes in Alaska. Am. J. Bot., 90: 1470-1480.
Direct Link - Cordell, S., G. Goldstein, D. Mueller-Dombois, D. Webb and P.M. Vitousek, 1998. Physiological and morphological variation in Metrosideros polymorpha, a dominant Hawaiian trees species, along an altitudinal gradient: The role of phenotypic plasticity. Oecologia, 133: 188-193.
Direct Link - De la Vega, M.P., 1996. Plant genetic adaptedness to climatic and edaphic environment. Euphytica, 92: 27-38.
CrossRefDirect Link - Struwe, L., V.A. Albert and B. Bremer, 1994. Cladistics and family level classification of the Gentiales. Cladistics, 10: 175-206.
Direct Link - Ferguson, I.K., B.D. Schrire and R. Shepperson, 1994. Pollen Morphology of the Tribe Sophoreae and Relationships Between Subfamilies Caesalpinoideae and Papilionoideae. In: Advances in Legume Systematics: Part 6 Structural Botany, Ferguson, I.K. and S.C. Turker (Eds.). Royal Botanic Garden, Kew, pp: 53-96.
- Erdtman, G., 1969. Handbook of Palynology: Morphology, Taxonomy, Ecology: An Introduction to the Study of Pollen Grains and Spores. Hafner, New York, Pages: 486.
Direct Link - Gottlieb, O.R., A.C. Kaplan and D.H.T. Zocher, 1993. A Chemosystematic Overview of Magnoliidae, Ranunculidae, Caryophyllidae and Hamamelidae. In: The Families and Genera of Vascular Plants, Flowering Plants Dicotyledons: Magnoliid, Hamamelid and Caryophyllid Families, Kubitzki, K., J.G. Rohwer and V. Bittrich (Eds.). Vol. II, Springer-Verlag, Berlin, pp: 20-31.
- Jensen, U., 1995. Secondary compounds of the Ranunculiflorae. Plant Syst. Evol. Suppl., 9: 85-98.
Direct Link - Erbar, C. and P. Leins, 1994. Flowers in Magnoliidae and the Origin of Flowers in other Subclasses on the Angiosperms, I. The Relationships Between Flowers of Magnoliidae and Alismatidae. In: Endress, P.K. and E.M. Fiiris (Eds.), Early Evolution of Flowers, Endress, P.K. and E.M. Fiiris (Eds.). Springer-Verlag, Vienna, pp: 193-208.
- Baroux, C., C. Spillane and U. Grossniklaus, 2002. Evolutionary origin of the endosperm in flowering plants. Genome Biol., 3: 1026.1-1026.5.
Direct Link - Barthlott,W. and I. Theisen, 1995. Epicuticular wax ultrstructure and classification of Ranunculiflorae. Plant Syst. Evol. Suppl., 9: 39-45.
Direct Link - Kadereit, J.W., F.R. Blattner, K.B. Jork and A. Schwarzbach, 1995. The Phylogeny of the Papaveraceae Senso Lato: Morphological, Geographical and Ecological Implications. In: Systematics and Evolution of the Ranunculiflorae, Jensen, U. and J.W. Kadereit (Eds.). Springer-Verlag, Vienna, New York, pp: 133-145.