
Mohd Razi Ismail
Not Available
Mohd Kamil Yusoff
Not Available
Marziah Mahmood
Not Available
Asian Journal of Plant Sciences
Year: 2004 | Volume: 3 | Issue: 6 | Page No.: 709-713







ABSTRACT
Young tissue cultured banana plants (Musa spp. Cv Pisang mas and Berangan) were subjected to different treatments of water deficit. Plants subjected to reduced water availability showed marked decreases in plant vegetative growth and perturbation in water relations and stomatal response. Stem girth and leaf elongation were markedly reduced after 7 days of withholding water. Regardless of a large depletion in soil moisture content, there was only a small reduction in leaf water status measured as relative water content. Stomatal conductance was more sensitive to the depletion of soil moisture content compared to leaf water status. A large and early decrease in stomatal conductance was shown not to be correlated with the reduction in relative water content. Proline concentration in banana leaf tissue was increased with the onset of water stress. The study also showed that stomatal conductance of banana plants was affected by the changes in Vapor Pressure Deficit (VPD) in both irrigated and water-stressed plants.
PDF Abstract XML References Citation
How to cite this article
Mohd Razi Ismail, Mohd Kamil Yusoff and Marziah Mahmood, 2004. Growth, Water Relations, Stomatal Conductance and Proline Concentration in
Water Stressed Banana (Musa spp.) Plants. Asian Journal of Plant Sciences, 3: 709-713.
DOI: 10.3923/ajps.2004.709.713
URL: https://scialert.net/abstract/?doi=ajps.2004.709.713
DOI: 10.3923/ajps.2004.709.713
URL: https://scialert.net/abstract/?doi=ajps.2004.709.713
INTRODUCTION
Water deficit is one of the principal environmental limiting factors for crop production throughout the world. Water deficit, which is a consequence of either intermittent or terminal period of drought, causes significant yield reduction on presently cultivated lands. These restrictions on yield potential are of great concern in terms of meeting food demand for accelerated increasing world population. The problem becomes more complicated by the fact that supplies of suitable irrigation water are dwindling rapidly and cost of irrigation are becoming prohibitive. Therefore, deliberate efforts are needed for better water shed management incorporating integrated approaches to water conservation. To achieve these objectives, understanding the fundamentals of plant responses is vital for a given species of crop within a specific environment.
Banana cultivation has been practiced either in small or large commercial scale in Malaysia. The plants require uniformly warm and moist conditions for optimum growth and yield. The need to provide adequate water supply for growth and plant development is well-documented[1,2]. Depletion of soil water from the root zone affects many plant physiological and biochemical processes[3]. In Malaysia, studies on the relations of banana plant and water supply are scarce due to an assumption that rain-fed water supply is already adequate for growth and development of banana. The present investigation was undertaken to determine the effects of water deficit on the growth and physiological processes of banana plants.
MATERIALS AND METHODS
Experiments were conducted both in the protected structure and under field conditions. In the first experiment, three month-old tissue cultured banana plants Cv Pisang mas were grown in soil (Munchong Series; Tropeptic Harplorthox) in 55x55x45 cm size containers under protected structure at Field 10B, Universiti Putra Malaysia. Plants were divided into two groups with well watered and withholding water. Watering was done to the soil maintain moisture at field capacity for well watered condition. For water stressed plants, water was with held after one last watering at field capacity. Soil moisture content after with holding water is shown in Fig. 1.
In another experiment, banana Cv. Berangan plants were grown under field condition at the Integrated Fruit Cultivation Project of Department of Agriculture and Department of Irrigation and Drainage in Tanah Merah, Kelantan. Plants were either irrigated or not irrigated (rainfed). Water for irrigated plot was supplied through a drip irrigation system.
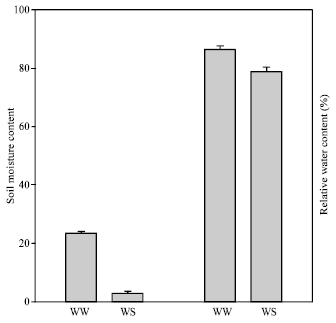 | |
| Fig. 1: | Soil moisture content and relative water content measured on the 20th day of water stressed. WW= well watered; WS: water stressed |
Each individual plant was supplied with two drippers adequate to wet both sides of the plant.
In the first experiment, stem height, diameter and elongation of leaf were recorded at 3-4 days interval throughout the drying period lasting for 20 days. At harvest, leaf area and dry weight of leaf, stem and root were determined.
Three to four young fully expanded leaves were chosen from each treatment for the determination of stomatal resistance and leaf photosynthetic rate. Stomatal resistance was taken with a transit-time prometer (AP-4, Delta-T, Cambridge, UK) calibrated with a perforated plate of known resistance. A portable open system infrared gas analyzer (LCA3) was used to determine the leaf photosynthetic rate. Relative water content was determined following the procedure described by Turner[4]. Leaves adjacent to the leaves sampled for relative water content were taken for the determination of proline accumulation. Determination of proline was carried out as described by Bates et al.[5]. A 0.5 g quantity of leaf was ground in 10 mL of 3% sulphosalicyclic acid and the extract filtered through a Whatman No. 2 filter paper. Aliquoats of 2 mL were used for proline estimation by the acid ninhydrin method and the reading recorded using a spectrophotometer (Shimadzu UV-160A Visible Recording Spectrophotometer, Japan).
In the second experiment, seasonal variations of stomatal conductance and soil moisture content were recorded. Diurnal variation of stomatal conductance and the changes in Vapor Pressure Deficit (VPD) were recorded on several occasions. VPD is calculated form an equation, VPD = Es (1-RH/100) where, Es (Pa) is the saturation water vapour of air derived form a psychrometric table.
RESULTS AND DISCUSSION
In the present study, leaf number of the water stressed plants was significantly lower compared to that of control plants (Fig. 2). This effect was apparent at day 10 from the onset of the water stress while the rate of new leaf appearance remained constant up to day 20. The same trend was observed in the leaf elongation rate but the severity of the effect started 7 days from the imposition of water stress after which no further growth increment was observed. This probably indicates that leaf growth rate is more sensitive to water stress than the rate of new leaf appearance although both parameters have been affected. Hoffmann and Turner[6] indicated that the ratio of leaf emergence does not appear to be a suitable indicator for plant response to water deficit.
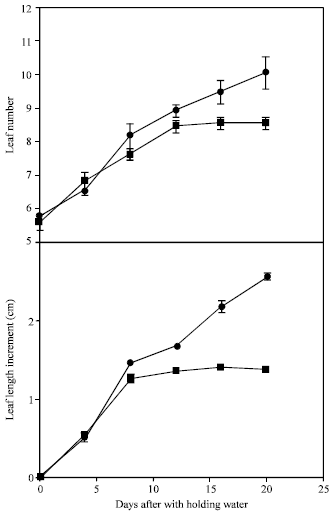 | |
| Fig. 2: | Leaf number and length increment of young banana plants grown under (●) well watered and (■) water stressed conditions. Bars represent ±SE means of 5 replicates |
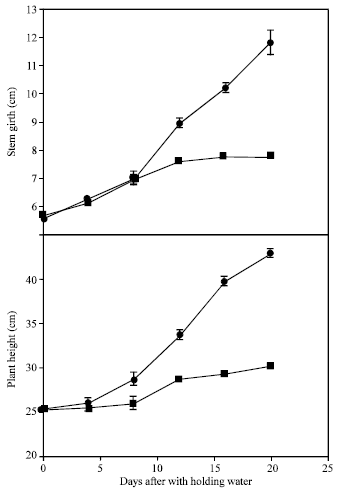 | |
| Fig. 3: | Stem girth and height of young banana plants grown under (●) well watered and (■) water stressed conditions. Bars represent ±SE means of 4 replicates |
A progressive increase in plant height was obvious in well-watered plants with time (Fig. 3A). Water stress treatment strongly reduced the height of plants. The rate of expansion growth was significantly lower from day 7 after which plants exhibited slight increment up to day 10 and no increase was observed thereafter, resulting in stunted plants as compared with the well-watered ones. The patterns of stem girth expansion growth were very similar to that of plant height. The most evident effect of water deficit to the plants was growth inhibition[7]. Cell enlargement is one of the most sensitive process affected by a change in plant water status[8]. It is a common phenomenon that pseudostem size increased progressively with plantation age when grown under optimum conditions, hence it is true for well-watered plants, but the current results indicate the severity of the stress on the growth rate during the early stage of plantation establishment. Besset et al.[9] showed a similar respond of stem expansion when water stress was applied at the final stage of rapid growth peach trees.
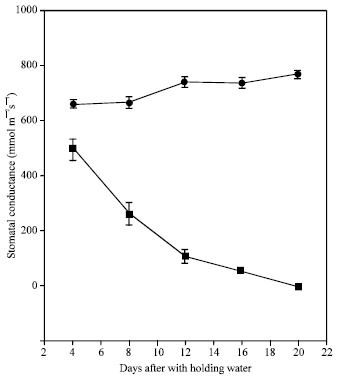 | |
| Fig. 4: | Changes in stomatal conductance of banana plants grown in (●) well watered and (■) water stressed conditions. Bars represent SE means of 5 replicates |
Ortuno et al.[10] showed that the first detectable response to the deficit irrigation was a marked increase in maximum daily trunk diameter shrinkage followed by a decrease in sap flow, predawn leaf water potential and conductance values in lemon trees.
Stomatal conductance was sensitive to the onset of water deficit (Fig. 4 and 5). The stomatal conductance of stressed plants grown under protected structure, started to decline as soon as the water stress was imposed (after 2 days) and rapid decline continued up to day 20 where it displayed complete stomatal closure. For water stressed plants under field conditions, stomatal conductance started to decline at the onset of water stress treatments. (Fig. 5A) and remained generally lower than the control plants throughout the experiment. The marked reduction in stomatal conductance of plants grown in the field was clearly observed when measurements were taken during severe drought stress at the location (Apr-July 1997). The decline in stomatal conductance was associated with the soil moisture content (Fig. 5C) and VPD (Fig. 6) and not with plant relative water content (Fig. 5B), indicating that the VPD-driven transfer of water vapor from the leaf was becoming limited by stomatal closure. These results are consistent with observation on oil palm, whereby stomatal conductance was reduced even though plants were grown in adequate soil moisture[11].
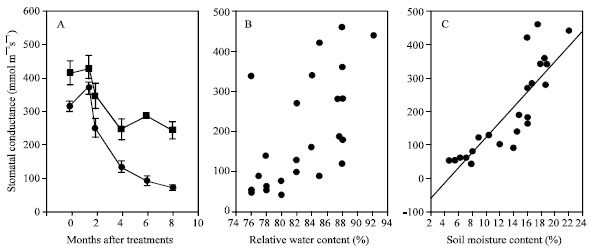 | |
| Fig. 5: | Water relations of banana plants ; A: Changes in stomatal conductance under field conditions■; well watered; ●=water stressed. B: Relationship between relative water content and stomatal conductance C: Relationship between stomatal conductance and soil moisture content |
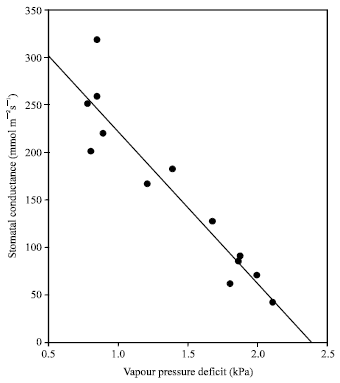 | |
| Fig. 6: | Relationships between Vapour Pressure Deficit and stomatal conductance of plants grown under field conditions (n= 15) |
The early reduction in stomatal conductance could also be due to reduced soil or root water potential rather than that of relative water content (Fig. 5C). This experiment under field conditions revealed that the physiological constraints due to adverse climatic conditions which prevailed during the growth of banana plants caused abnormal plant development where fruit bearing was delayed for at least 4 months. Furthermore, the productivity of banana plants at the experimental plot was severely reduced compared to the usual yield of the varieties grown elsewhere in Malaysia.
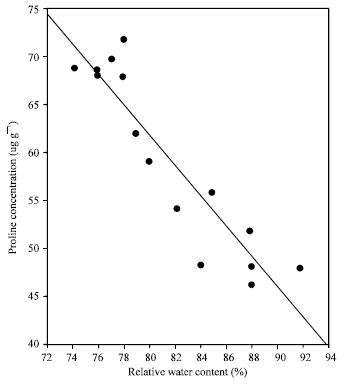 | |
| Fig. 7: | Relationships between proline and relative water content in banana leaves (n= 15) |
This observations strongly suggest that water is a primary factor governing the growth and yield of banana.
Level of proline accumulation in banana leaf tissue was associated with the internal water status of the plant. The proline levels started to increase from 45 ug g-1 to a value of 74 ug g-1 fresh weight when the relative water content of the leaf tissue started to decrease with the onset of water stress from 92 to 72%, in both sites of the experiment (Fig. 7). It is widely accepted that water deficit enhances accumulation of proline in many plant species and that proline may regulate the osmotic balance of the cell thus relieving the negative effect of stress[12-14].
This study suggests that water management in banana cultivation is vital for its growth and development. There is a need to determine strategies to improve efficient water use of plants during drought conditions. Further studies should emphasise on the water requirement of banana plants at different growth stages to determine the sensitivity of growth phases to water stress.
ACKNOWLEDGMENT
We wish to thank Research Grant No. 01-02-04-0518-EA001 from Ministry of Science, Technology and Environment, Malaysia (MOSTE) for funding this project.
REFERENCES
- Turner, N.C., 1981. Techniques and experimental approaches for the measurement of plant water status. Plant Soil, 58: 339-366.
CrossRefDirect Link - Bates, L.S., R.P. Waldren and I.D. Teare, 1973. Rapid determination of free proline for water-stress studies. Plant Soil, 39: 205-207.
CrossRefDirect Link - Hoffmann, H.F. and D.W. Turner, 1993. Soil water deficits reduce the elongation rate of emerging banana leaves but the night/day elongation ratio remains unchanged. Sci. Hortic., 54: 1-12.
Direct Link - Hsiao, T.C. and E. Acevedo, 1974. Plant responses to water deficits, water-use efficiency and drought resistance. Agric. Meteorol., 14: 59-84.
CrossRefDirect Link