
S. Aghighi
Not Available
G.H. Shahidi Bonjar
Not Available
R. Rawashdeh
Not Available
S. Batayneh
Not Available
I. Saadoun
Not Available
Asian Journal of Plant Sciences
Year: 2004 | Volume: 3 | Issue: 4 | Page No.: 463-471







ABSTRACT
This is the first report of antifungal activity of Iranian Actinomycetes isolates against Alternaria solani Sorauer, Alternaria alternata (Fr.) Keissler, Fusarium solani (Mart.) Sacc., Phytophthora megasperma Drechsler, Verticillium dahliae Klebahn and Saccharomyces cerevisiae Meyen ex Hansen. Biological control of plant diseases has received worldwide attention in recent years mainly as a response to public concern about the use of hazardous chemicals in the environment. Soil Actinomycetes particularly Streptomyces spp. enhance soil fertility and have antagonistic activity against wide range of soil-borne plant pathogens. In search for soil Actinomycetes having antifungal activity against plant fungal-pathogens, 110 isolates were screened from which 14 isolates were found active at least against one of the tested fungi. Streptomyces plicatus strain101, Frankia sp. strain 103 and Streptomyces sp. strain 44 had the widest antifungal spectra of activity. Antifungal genes from these strains may be proper candidates for genetic engineering of plants for increased tolerance against the tested cosmopolitan fungal pathogens.
PDF Abstract XML References Citation
How to cite this article
S. Aghighi, G.H. Shahidi Bonjar, R. Rawashdeh, S. Batayneh and I. Saadoun, 2004. First Report of Antifungal Spectra of Activity of Iranian Actinomycetes Strains Against Alternaria solani, Alternaria alternate, Fusarium solani, Phytophthora megasperma, Verticillium dahliae and Saccharomyces cerevisiae. Asian Journal of Plant Sciences, 3: 463-471.
DOI: 10.3923/ajps.2004.463.471
URL: https://scialert.net/abstract/?doi=ajps.2004.463.471
DOI: 10.3923/ajps.2004.463.471
URL: https://scialert.net/abstract/?doi=ajps.2004.463.471
INTRODUCTION
Saprophytic Gram-positive bacteria in the genus Streptomyces are the most widely studied and well known genus of the Actinomycete family. Streptomycetes usually inhabit soil and are important decomposers. They are able to metabolize many different compounds including sugars, alcohols, amino acids and aromatic compounds by producing extracellular hydrolytic enzymes. Their metabolic diversity is due to their extremely large genome which has hundreds of transcription factors that control gene expression, allowing them to respond to specific needs[1]. Streptomycetes are also of medical and industrial importance because they synthesize antibiotics. There are several theories to may explain antibiotic production; the most widely accepted one being that antimicrobial compounds help the organism compete with other organisms in the relatively nutrient-depleted environment of the soil by reducing competition. Over 50 main antibiotics have been isolated from Streptomycetes species, including streptomycin, neomycin, chloramphenicol and tetracyclines[2].
In modern agriculture, pesticide application is still an invaluable and effective method to control plant diseases. However, since use of agrochemicals is falling into disfavor because of environmental pollution and detrimental effects on a variety of nontarget organisms, potential use of microbes based biocontrol agents as replacement or supplements for agrochemicals has been addressed in many recent reports[3]. With the increased concern about conserving natural resources as air, soil and water, natural or biological control of plant diseases has received increased emphasis. Biological control of plant diseases is slow, gives few quick profits, but can be long lasting, inexpensive and harmless to life. Biocontrol systems do not eliminate neither pathogen nor disease but bring them into natural balance[4]. In search for biocontrol agents, several fungi as Talaromyces flavus, Gliocladium spp., Trichoderma spp., Aspergillus giganteus and Penicillium chrysogenum and bacteria as Bacillus subtilis, Pseudomonas spp. and Actinomycetes (especially Streptomyces spp.) have shown activity against several plant pathogens[5-11].
Streptomyces spp. have been shown to have characteristics which make them useful as biocontrol agents against soil-borne fungal plant pathogens. These characteristics include the production of different kinds of secondary metabolites and biologically active substances of high commercial value such as enzymes (which degrade the fungal cell wall directly) and antibiotics and they are of the major contributors to the biological buffering of soils and have roles in decomposition of organic matter conductive to crop production[12-17]. Several workers have reported that in vitro studies have documented satisfactory results in use of Streptomyces against some root pathogens. For example, Streptomyces sp. strain 5406 has been used in China for the last 35 years to protect cotton crops against soil-borne pathogens[18]. The results even show that use of Streptomyces enhances growth of the crops and vegetables[19,20]. Some of commercially produced antifungals driven from Streptomyces are: Cycloheximide from S. griseus used in control of bacterial diseases of lawn and cherry leaf spot; Blastcidin-S from S. griseochromogenes and Kasugamycine from S. kasugaensis used in control of rice blast pathogen[21] Rhizovit® from S. rimosus and Mycostop® from S. griseoviridis that are used in control of wide range of fungi as Pythium spp., Fusarium spp., Phomopsis sp., Phytophthora spp. Rhizoctonia solani, Alternaria brassicola, Botrytis sp. and Fusarium spp.[22,23]. Some workers have reported biological control of Verticillium by Streptomyces spp. Chi and Hanson[24] reported in vitro antifungal activity of S. rimosus against V. dahliae and V. albo-atrum. There are also reports on control of root fungal-pathogens by use of seed and seedling treatments with spore suspensions of some Streptomyces isolates[25].
Studies on biological control of mentioned phytopathogenic fungi have a long history because available control methods are either inefficient or difficult to apply[26]. With extended environmental diversity, however, the Actinomycetes microflora of the Iranian soils has not been very well explored with the goal of exploring new means of biocontrols. With the respect to their role in biological control of soil-borne fungal-pathogens, at the present research 110 isolates of Actinomycetes were isolated from agricultural soils of Kerman province, Iran and screened against Alternaria solani, Alternaria alternata, Fusarium solani, Phytophthora megasperma and Verticillium dahliae. These are ubiquitous and cosmopolitan phytopathogens causing severe diseases in wide range of crops[21,27,28]. For better evaluation of the spectral activity of the isolated Actinomycetes, the yeast Saccharomyces cerevisiae was also included in the survey. It is commonly known as brewer's yeast, but has also been reported as the cause of both superficial and invasive infections in humans[29]. From 110 isolates of Actinomycetes, 14 isolates showed antifungal activity at least against one of the tested fungi.
MATERIALS AND METHODS
Culture media: A synthetic medium, Casein glycerol (or starch) agar (CGA) was used for screening and isolating of Actinomycetes which composed of: glycerol or soluble starch, 10 g; casein, 0.3 g; KNO3, 2 g; NaCl, 2 g; K2HPO4, 2 g; MgSO4.7H2O, 0.05 g; CaCO3, 0.02 g; FeSO4.7H2O, 0.01 g and agar, 18 g in 1 L of distilled H2O (pH 7.2)[4]. In submerged cultures, Agar was excluded (CG medium). Actinomycetes colonies with different morphologies were selected and transferred to CGA slants for further studies[30,31].
Preparation of fungal isolates: Tested fungi were kind gifts as follows: Alternaria solani (tomato isolate) from Dr. Zare, Mycology Lab., Plant Pests and Diseases Research Institute, Evin, Tehran; Alternaria alternata (tomato isolate) from Mycology Lab., Department of Plant Pathology, College of Agricultural Sciences, Bahonar University of Kerman, Kerman; Fusarium solani (potato isolate) from Dr. Sharifnabi, Dept. of Plant Pathology, College of Agricultural Sciences, Industrial University of Isfahan, Verticillium dahliae (cotton, potato and olive isolates) from Prof. Banihashemi, Mycology Lab., Department of Plant Pathology, College of Agriculture, Shiraz University, Shiraz; Phytophthora megasperma and Verticillium dahliae (pistachio isolates) from Mr. Aminaii, Plant Pathology Lab., Agricultural Research Center, Ministry of Agriculture, Kerman, Iran. The fungi were grown at 25-26°C and maintained on potato dextrose agar (PDA) (Difco). Saccharomyces cerevisiae (PTCC No. 5052), obtained from the Persian Type Culture Collection, Tehran, Iran (PTCC) and grown at 30°C on Sabouraud dextrose medium. All cultures stored at 4°C and sub-cultured as needed.
Isolation of actinomycetes from soil: Soil samples were collected from grasslands, orchards and vegetable fields in different localities of Kerman province, Iran. Several samples randomly were selected from mentioned localities using an open-end soil borer (20 cm indepth, 2.5 cm in diameter) as described by Lee and Hwang[12]. Soil samples were taken from a depth of 10-20 cm below the soil surface. The soil of the top region (10 cm from the surface) was excluded. Samples were air-dried at room temperature for 7-10 days and then passed through a 0.8 mm mesh sieve and were preserved in polyethylene bags at room temperature before use. Samples (10 g) of air-dried soil were mixed with sterile distilled water (100 ml). The mixtures were shaken vigorously for 1 h and then allowed to settle for 1 h. Portions (1 ml) of soil suspensions (diluted 10-1) were transferred to 9 ml of sterile distilled water and subsequently diluted to 10-2,10-3 ,10-4 ,10-5 and 10-6. Inocula consisted of adding aliquots of 10-3 to 10-6 soil dilutions to autoclaved CGA (1 ml -25 ml CGA) at 50°C before pouring the plates and solidification. Three replicates were considered for each dilution. Plates were incubated at 30°C for up to 20 days. From day 7 on, Actinomycetes colonies were isolated on CGA, incubated at 28°C for one week and stored refrigerated as pure cultures before use. For screening studies 110 pure Actinomycetes isolates were collected.
Screening procedures and in vitro antifungal bioassays
Agar disk-method: Each Actinomycetes isolate was smeared on CGA medium as a single streak and after incubation at 28°C for 4-6 days, from well-grown streaks 6 mm Agar disks of Actinomycetes colony mass was prepared by using sterile cork borers. Disks were then aseptically transferred to PDA plates having fresh lawn cultures of fungal isolates. Controls included using plain disks from CGA medium. Plates were incubated at 25-26°C for 4-6 days and bioactivity was evaluated by measuring the diameter of inhibition zones (DIZ, mm)[4,32].
Dual culture bioassay: Fungal mycelial-disks (diameter of 6mm) prepared from growing margin of cultures of test fungal isolates and placed in the center of PDA plates and at 30 mm distance from it, the Actinomycetes disks (prepared as mentioned) were placed. Plates incubated at 27°C for 2-3 weeks[4]. Antifungal activity was indicative as mycelial growth of fungal isolates was prohibited in the direction of active Actinomycetes isolate. The level of inhibition at dual cultures was calculated by subtracting the distance (mm) of fungal growth in the direction of an antagonist colony (γ) from the fungal growth radius (γB) of a control culture to give Δγ = γB-γ. The ratings used were modified from those of Lee et al.[12] and El-Tarabily et al.[33]. where, Δγ: 5-9 mm, + (weak inhibition); Δγ: 10-19 mm, ++ (moderate inhibition) and Δγ>20 mm, +++ (strong inhibition). Controls included fungal mycelial plugs in center of non-Actinomycetes inoculated PDA plates.
Agar fungal-strip method: From well grown fungal cultures 1x7 cm strip was cut aseptically and used as described above in dual culture bioassay.
Well diffusion-method: For evaluation of antifungal activity of aqueous samples, by use of sterile cork borer wells (6x4 mm, 2 cm apart) were punctured in fresh fungal lawn-cultures or at 30 mm distance from fungal plugs. Respective concentrations in dimethyl sulfoxide: methanol (1/1: v/v) solvent (DM solvent) were then administered to fullness in each well. Plates were incubated at 26°C for 4-6 days for lawn cultures and 14 days for dual culture disk-plugs. Bioactivity was determined by measuring inhibitory zones (mm). Each experiment was repeated three times and the mean of inhibitory zones recorded. Controls included use of blank wells and use of DM solvent without test compounds[4].
Preparation of crude extract from submerged cultures: Active strains of Actinomycetes were grown in submerged cultures of CG medium on rotary shakers under 130 rpm at 30°C. To monitor the activity, aseptically small aliquots of culture media were taken every 24 h for 20 days and the activity was evaluated by well diffusion-method[4,32]. To prepare crude extracts, depend to Actinomycetes isolate, between 9 to 12th days of post inoculation which the activity reached maximum, the cultures were harvested; spores and mycelia were excluded by filtration through two layers of cheese cloth. The clarified sap was then dried to dark crude under reduced air at 50°C and kept refrigerated before use.
Classification of actinomycetes: Actinomycetes colonies were characterized morphologically and physiologically to the genus level following the direction mentioned in the methods manual of international cooperative project for description and deposition of cultures of Streptomyces (ISP)[34].
Morphological characterization: Actinomycetes colonies on glycerol-nitrate-casein agar were transferred onto oatmeal agar and streaked across the plate and incubated in the dark at 27°C for 21 days.
Color determination: This made for: a) Mass color or mature, sporulating aerial surface growth, b) The color of substrate mycelium as viewed from the reverse side and c) Diffusible soluble pigments other than melanin. Observation was made after 21 days and was limited to mature cultures with heavy spore mass surface using code for determining the color of aerial mycelium of Streptomycetes composed by Prauser[35] for color tabs of Baumann Farbtonkarte Atlas I.
Determination of morphological characteristic of the sporebearing hyphae: The spore-bearing hyphae characteristics were determined by direct microscopic examination of the culture surface (21 days old) on opened dishes of the crosshatched cultures using 100 x magnification. The species involved in the genus Streptomyces divide into sections: Rectus (R) or straight, flexible (F) or flexeous, Retinaculum-Apertum (RA) and spiral (S).
Melanin production: Peptone iron agar was used for the detection of deep brown to black diffusible pigment (+). Absence of the color was recorded as negative (-).
Carbon utilization: The following sugars were tested, L-arabinose, D-xylose, meso-inositol, D-mannitol, D-fructose, rhamnose, raffinose and sucrose. Preparation was done as described in the ISP[34].
RESULTS AND DISCUSSION
Screening and bioassays: In screening for Actinomycetes having antifungal activity, 110 isolates were screened from which 14 isolates showed activity against at least one fungal isolate. Table 1 shows antifungal activity of active Actinomycetes strains on eight phytopathogenic fungal isolates and Saccharomyces cerevisiae. Colony morphology of Streptomyces plicatus Strain 101 (A), Streptomyces sp. Strain 44 (B) and Frankia sp. Strain 103C at 40 X magnification is shown in Fig. 1.
Antifungal reaction types: As revealed by microscopic examinations, antagonistic activity of S. plicatus strain 101 against isolates of V. dahliae consists of two inhibitory effects. Clear zones adjacent to the wells which represent complete mycelial growth (fungicidal activity) and in their periphery, there is microsclerotia inhibition zone (Fig. 2A) which indicates inhibition of melanin formation in the pathogen. Inhibitory effect of Frankia sp. Strain 103 consisted mainly of the latter type (Fig. 6B) against V. dahliae. Except for V. dahliae, other tested fungi represented clear inhibition zones (Fig. 2-7).
Taxonomy of actinomycetes: Strains 101 and 103 were identified as Streptomyces plicatus[36] and Frankia sp., respectively. Based on the literature reviews; both are new records from Iran.
Actinomycetes produce more than half of the world's antimicrobials and are consequently becoming valuable tools in the field of biological control. Antifungal activity of the isolates found in this study highlights their importance as candidates for further investigation in biological control of tested pathogens. Broad spectral activity of strains 101, 103 and 44 are more noticeable. Phytophthora megasperma was the most and Saccharomyces cerevisiae and Fusarium solani were the least sensitive fungi, respectively.
| Table 1: | Antifungal inhibitory effect of active Actinomycetes strains against eight phytopathogenic fungal isolates and Saccharomyces cerevisiae. |
 | |
| a: Alternaria solani (tomato isolate); b: Alternaria alternata (tomato isolate); c: Fusarium solani (potato isolate); d: Phytophthora megasperma (pistachio isolate); e: Verticillium dahliae (pistachio isolate); f: Verticillium dahliae (cotton isolate); g: Verticillium dahliae (potato isolate); h: Verticillium dahliae (olive isolate); I: Saccharomyces cerevisiae (PTCC No. 5052). +: 5-9 mm, weak; + +: 10-19 mm, moderate; +++: > 20 mm, strong and-: no inhibition. *: Inhibition of microsclerotia and melanin production in V. dahliae isolates | |
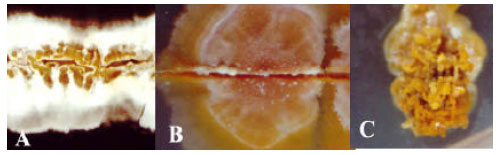 | |
| Fig. 1: | Aerial colony mats of Streptomyces plicatus Strain 101 (A), Streptomyces sp. Strain 44 (B) and Frankia sp. Strain 103 © at 40 X magnification |
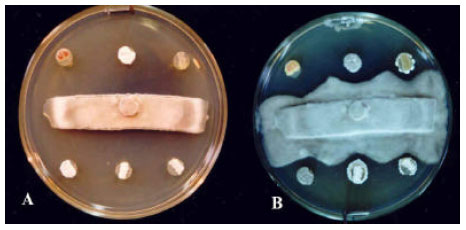 | |
| Fig. 2: | Antifungal activity of 6 isolates of Actinomycetes against Alternaria solani indicated by Agar fungal-strip method. A) Clockwise from top left: Streptomyces sp. Strain 44, S. plicatus Strain 101, Frankia sp. Strain 103, Streptomyces spp. Strains of 102, 56 and 78 at the start of bioassay and B) after 10 days of incubation, respectively |
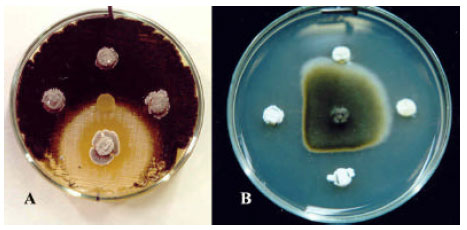 | |
| Fig. 3: | Antifungal activity of Streptomyces isolates indicated by Agar disk method against A) lawn culture of pistachio root-isolate of Verticillium dahliae, Clockwise from bottom: S. plicatus Strain 101, Streptomyces spp. Strains of 102, 01 and N. Middle represents blank agar disk (control); B) Alternaria alternata, clockwise from top: Streptomyces spp. Strains of 43, 95, 102 and S. plicatus Strain 101. Middle is A. alternata inoculation disk |
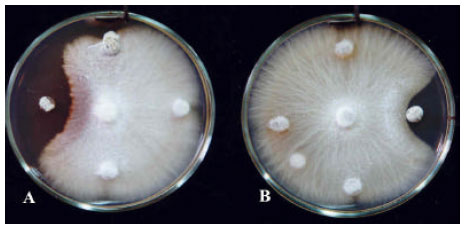 | |
| Fig. 4: | Antifungal activity of Streptomyces isolates indicated by Duel culture-method against Fusarium solani, Clockwise from tops: A) strains 96, 78, 30 and S. plicatus Strain 101; B) strains 56, 102, 01, N and G. Middles are F. solani inoculation disks |
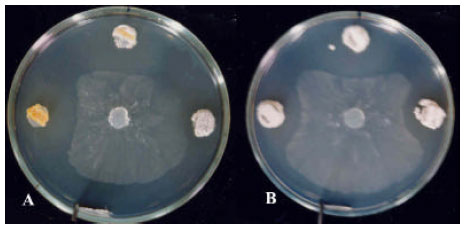 | |
| Fig. 5: | Antifungal activity of Actinomycetes isolates indicated by Duel culture-method against pistachio root-isolate of Phythophthora megasperma, Clockwise from tops: A) Frankia sp. Strain 103, Streptomyces spp. Strains of 01 and 44; B) Streptomyces spp. Strains of L, N and 30 |
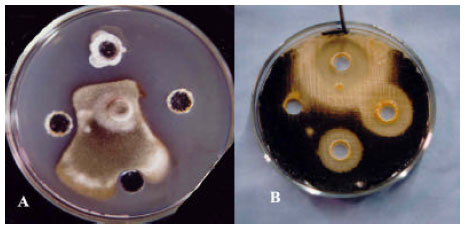 | |
| Fig. 6: | Antifungal activity of culture filtrate of Actinomycetes isolates in well-diffusion method against A) Alternaria alternatA, Clockwise from top: Streptomyces sp. Strain 102, S. plicatus Strain 101, Streptomyces spp. Strains of 30 and L; B) Lawn culture of pistachio root-isolate of Verticillium dahliae, lockwise from top: S. plicatus Strain 101, Streptomyces sp. Strain 44, Frankia sp. Strain 103 and control well (uninoculated liquid medium) |
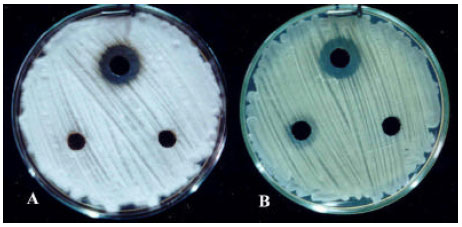 | |
| Fig. 7: | Antifungal activity of culture filtrate of Streptomyces isolates in well-diffusion method against lawn culture of A) Fusarium solani, Clockwise from top: S. plicatus Strain 101, Streptomyces spp. Strains of 43 and 95;) Saccharomyces cerevisiae, Clockwise from top: S. plicatus Strain 101, Streptomyces spp. Strains of 96 and 78 |
Control of root pathogens has largely been based on soil fumigation with broad-spectrum biocides, most of which will soon be banned from the market because of their adverse effects on the environment. With this worldwide perspective, to help the soil ecosystem environmentally safe and non chemical-measures are to be developed for combating plant diseases. In ideal biological control measures, proper microorganisms are those having well adaptation in soil and rhizosphere exerting effective antagonistic activity against soil pathogens persistently. In this regard soil-driven Actinomycetes do not have adverse effect or alter the biological buffering of soils as chemical measures do. They should receive higher attention in research for biological controls worldwide.
Phytopathogenic fungi are of major problems in agriculture. Genetic engineering provides an opportunity to protect plants from fungal diseases and to reduce the use of synthetic fungicides. The genes for antifungal metabolites can be engineered into plants to increase the resistance of crop plants to fungal attack, decreasing the use of environmentally unfriendly fungicides. The major factor limiting the application of this technology is the identification and isolation of useful genes that code for antifungal metabolites. Plants themselves are a potential source of new antifungal activities. For example, two antifungal proteins have been isolated from radish. The genes for these proteins were cloned and constitutively expressed in transgenic plants, which showed enhanced resistance to pathogenic fungi[37, 38].
The main objective in our study was to identify biologically active Actinomycetes isolates against the cosmopolitan plant pathogens. Preliminary data in this paper describes the presence of potential antifungal metabolite(s) in Streptomyces plicatus (strain101), Frankia sp. (strain103) and Streptomyces sp. strain 44 against tested fungi. These strains prohibit mycelial growth, formation of microsclerotia and melanin production in Verticillium dahliae. Except for V. dahliae, other tested fungi represented clear inhibition zones.
Lack of melanin renders the pathogen more susceptible to antagonists and hence favors its biological control in soil.
Expression of cloned genes in transgenic plants has provided evidence in plant defense[13]. The genes encoding many antifungal proteins are currently being used by agribusiness to create genetically modified plants that have increased fungal resistance in field[39]. Expression of cloned chitinase genes in transgenic plants has provided evidence of their role in plant defense[40]. Results of present findings may form the avenue for production of resistant transgenic-plants with recombinant DNA having antifungal genes cloned from biologically active Actinomycetes. Thus it may be assumed that the antifungal-metabolite genes from Streptomyces plicatus Strain 101, Frankia sp. (strain103) and Streptomyces sp. strain 44 may be useful candidates for genetic engineering of agriculturally important crop plants for increased tolerance against tested phytopathogens. Having special environmental characteristics and being rich in Actinomycetes population, the microbiology of the Iranian soils has to be further explored for new active isolates of Actinomycetes.
ACKNOWLEDGMENTS
Thanks to Head of Research Affairs, Office of Bahonar University of Kerman for financial support of the project. Helpful information and kind gifts of fungi by Dr. Zare, Dr. Sharifnabi, Mr. Aminaii and Prof. Banihashemi are appreciated. This research is dedicated to Mr. A. Afzalipour, the founder of Bahonar University in Kerman.
REFERENCES
- Goshi, K., T. Uchida, A. Lezhava, M. Yamasaki, K. Hiratsu, H. Shinkawa and H. Kinashi, 2002. Cloning and analysis of the telomere and terminal inverted repeat of the linear chromosome of Streptomyces griseus. J. Bacteriol., 184: 3411-3415.
Direct Link - Shimizu, M., Y. Nakagawa, Y. Sato, T. Furumai and Y. Igarashi et al., 2000. Studies on endophytic actinomycetes (I) Streptomyces sp. isolated from Rhododendron and its antifungal activity. J. Genet. Plant Pathol., 66: 360-366.
CrossRefDirect Link - De Boer, M., P. Bom, F. Kindt, J.J.B. Keurentjes, I. van der Sluis, L.C. van Loon and P.A.H.M. Bakker, 2003. Control of Fusarium wilt of radish by combining Pseudomonas putida strains that have different disease-suppressive mechanisms. Phytopathology, 93: 626-632.
CrossRefDirect Link - Kucuk, C. and M. Kivanc, 2003. Isolation of Trichoderma spp. and determination of their antifungal, biochemical and physiological features. Turk. J. Biol., 27: 247-253.
Direct Link - Moreno, A.B., A.M. Pozo, M. Borja and B.S. Segundo, 2003. Activity of antifungal protein from Aspergillus giganteus against Botrytis cinerea. Phytopathology, 93: 1344-1352.
Direct Link - Cattopadhyay, S.K. and B. Nandy, 1982. Inhibition of Helminthosporium oryzae and Alternaria solani by Streptomyces longisporus (Krasil' nikov) Waksman. Plant Soil, 69: 171-175.
Direct Link - Whipps, J.M., 2001. Microbial interactions and biocontrol in the rhizosphere. J. Exp. Bot., 52: 487-511.
PubMedDirect Link - Lee, J.Y. and B.K. Hwang, 2002. Diversity of antifungal actinomycetes in various vegetative soils of Korea. Can. J. Microbiol., 48: 407-417.
Direct Link - Kong, L.D., D.D. Tzeng and C.H. Yang, 2001. Generation of PCR-based DNA fragments for specific detection of Streptomyces saraceticus N45. Proc. Natl. Sci. Council Roc., 25: 119-127.
Direct Link - Valois, D., K. Fayad, T. Barasubiye, M. Garon, C. Dery, R. Brzezinski and C. Beaulieu, 1996. Glucanolytic actinomycetes antagonistic to Phytophthora fragariae var. rubi, the causal agent of raspberry root rot. Applied Environ Microbiol., 62: 1630-1635.
Direct Link - Merriman, P.R., R.D. Price, J.F. Kollmorgen, T. Piggott and E.H. Ridge, 1974. Effect of seed inoculation with Bacillus subtilis and Streptomyces griseus on the growth of cereals and carrots. Aust. J. Agric. Res., 25: 219-226.
CrossRefDirect Link - Brown, M.E., 1974. Seed and root bacterization. Annu. Rev. Phytopathol., 12: 181-197.
CrossRefDirect Link - Saadoun, I., F. Al-Momani, H. Malkawi and M.J. Mohammad, 1999. Isolation, identification and analysis of antibacterial activity of soil streptomycetes isolates from North Jordan. Microbios, 100: 41-46.
Direct Link - Saadoun, I. and R. Gharaibeh, 2002. The Streptomycetes flora of Jordan and its potential as a source of antibiotics active against antibiotic resistant Gram negative bacteria. World J. Microbiol. Biotechnol., 18: 465-470.
Direct Link - El-Tarabily, K.A., M.H. Soliman, A.H. Nassar, H.A. Al-Hassani, K. Sivasithamparam, F. McKenna and G.E. Hardy, 2000. Biological control of Sclerotinia minor using a chitinolytic bacterium and actinomycetes. Plant Pathol., 49: 573-583.
Direct Link - Shirling, E.B. and D. Gottlieb, 1966. Methods for characterization of Streptomyces species. Int. J. Syst. Evol. Microbiol., 16: 313-340.
CrossRefDirect Link - Aghighi, S., G.H.S. Bonjar and I. Saadoun, 2004. First report of antifungal properties of a new strain of Streptomyces plicatus (Strain 101) against four Iranian phytopathogenic isolates of Verticillium dahliae, a new horizon in biocontrol agents. Biotechnology, 3: 90-97.
CrossRefDirect Link - Brogue, K., I. Chet, M. Holliday, R. Cressman and P. Biddle et al., 1991. Transgenic plants with enhanced resistance to the fungal pathogen Rhizoctonia solani. Science, 254: 1194-1197.
CrossRefDirect Link - Selitrennikoff, C.P., 2001. Antifungal proteins. Applied Environ. Microbiol., 67: 2883-2894.
CrossRefPubMedDirect Link