
Bimal Kumar Pramanik
Not Available
Toshiyuki Matsui
Not Available
Haruo Suzuki
Not Available
Yusuke Kosugi
Not Available
Asian Journal of Plant Sciences
Year: 2004 | Volume: 3 | Issue: 4 | Page No.: 398-402







ABSTRACT
Changes in sucrose synthase (SS) and sucrose phosphate synthase (SPS) activities and sugar content during postharvest senescence in broccoli (Brassica oleracea L. cvs. `Hartland` and `Sairin`) were studied. The first change in color became apparent between 2 and 3 d but whole head yellowed after 5 d of storage. The SS activity increased gradually in all portions although with few fluctuations in branchlets of both cultivars at the end of the storage time. In case of SPS, there was no inclining or declining pattern of the activity in any portion of the two cultivars at the end of the storage period. In both cultivars, branchlets showed higher SS and SPS activity than florets. Sucrose content gradually decreased in both portions of the two cultivars with time. Fructose content was higher than glucose and sucrose in the florets as well as branchlets of both cultivars. There was a highly significant negative correlation observed between the SS activity and sucrose content in the florets and branchlets of both cultivars. There was no significant correlation between sugar contents and SPS activity in any portions of both cultivars.
PDF Abstract XML References Citation
How to cite this article
Bimal Kumar Pramanik, Toshiyuki Matsui, Haruo Suzuki and Yusuke Kosugi, 2004. Changes in Activities of Sucrose Synthase and Sucrose Phosphate Synthase and Sugar Content During Postharvest Senescence in Two Broccoli Cultivars. Asian Journal of Plant Sciences, 3: 398-402.
DOI: 10.3923/ajps.2004.398.402
URL: https://scialert.net/abstract/?doi=ajps.2004.398.402
DOI: 10.3923/ajps.2004.398.402
URL: https://scialert.net/abstract/?doi=ajps.2004.398.402
INTRODUCTION
Broccoli (Brassica oleracea L.) is a highly perishable vegetable subject to rapid color and quality deterioration during storage after harvest. The edible parts of broccoli are compact cluster of unopened flower buds (florets) and a tender portion of stem (branchlets). The physiology of broccoli changes rapidly after harvest. Within the first 6 h after harvest, the sucrose content of the florets dropped to less than half the level present at harvest[1]. However, when exogenous sucrose is supplied back to the plant, senescence symptom can be delayed and can also increase the longevity of broccoli[2,3]. Thus, sucrose could directly influence the postharvest senescence of broccoli. Therefore, it is important to elucidate the enzymes of sugar metabolism particularly during storage. In broccoli, sucrose is the primary form of translocated sugar and present in small amount, with the other two major soluble sugars, glucose and fructose, in approximately equal amount, while sucrose activity is high[3,4]. The cleavage of sucrose into glucose and fructose is performed by invertase or SS in the cleavage direction. On the other hand, sucrose is known to be synthesized by two metabolic pathways, one is through sucrose synthase (SS) which catalyzes the reversible reaction: UDP (uridine 5’-diphosphate)-glucose+ fructose→ sucrose+UDP[5] and other through sucrose phosphate synthase which catalyzes the reaction: UDP glucose+fructose-6-phosphate→sucrose phosphate activities may determine the sucrose concentration in the tissue.
During storage, acid invertase activity gradually increases with a concomitant decrease in sucrose that lead to rapid deterioration of harvested produce. Many studies have been reported on sugar metabolism such as of changes in the activity of acid invertase of broccoli during storage[3-6] and also SS and SPS during growth, development and maturation of fruits and vegetables[7-10]. Less work has been done on the changes of SS and SPS activities in broccoli during storage. Therefore, this study was conducted to investigate the influence of SS and SPS on the postharvest life of two broccoli cultivars stored at 20°C.
MATERIALS AND METHODS
Plant materials: Two broccoli cultivars ‘Sairin’ and ‘Hartland’ were harvested from a commercially grown crop and transported to the laboratory. Harvested broccoli heads were stored at 20OC for 5 days. After 24 h interval, the broccoli heads of each variety were taken out from storage and florets were shaved off a razor blade and placed in the individual bag and immediately stored at –30°C until use.
Color assessment: Color changes in broccoli heads were determined with a chromameter (Minolta CR-200), equipped with an 8 mm measuring head. The meter was calibrated using the manufacturer’s standard white plate. Color changes were quantified in the L, a, b color space. Hue angle (h° = tan-1 ( b/a ) when a>0 and b>0 or h°=180° + tan-1 ( b/a ) when a<0 and b>0 ) was calculated from the a and b values[11]. On each head, three readings were taken from different portions.
Enzyme extraction for SS and SPS: Approximately 5 g sample from each portion was mixed with 1% of PVPP and 1 g sea sand. The sample was then homogenized with 10 ml of 0.2 M K-P buffer (pH 7.8) containing 10 mM ascorbate, 15 mM MgCl2, 1 mM EDTA and 1 M DTT using a cooled mortar and pestle. The resulting homogenate was then filtered through four layer of cotton cloth and the filtrate was centrifuged at 11 000 x g for 20 min. The total supernatant was dialyzed with 40 times diluted 0.2 M K-P buffer (pH 7.8) for 16 h and the inner solution was used as the crude enzyme. All extraction procedures were carried out at 0-4°C.
Enzyme assays for SS and SPS: SS and SPS activities were assayed at 37°C by the method described by Hubbard[12] with slight modifications. Reaction mixtures (70 μl) contained 50 mM Hepes-NaOH buffer (pH 7.5), 15 mM MgCl2, 25 mM fructose-6-phosphate, 25 mM glucose-6-phosphate and 25 mM UDP-glucose. The mixture was incubated for 30 min. at 37°C and the reaction was terminated with the addition of 70 μl of 30% KOH. Enzyme blanks were terminated with KOH at 0 min. Tubes were kept at 100°C for 10 min to destroy any unreacted fructose or fructose-6-phosphate. After cooling, 2 ml of anthrone reagent (150 mg anthrone with 100 ml of 15% H2SO4) was added and incubated in a 40°C water bath for 15 min. After cooling, color development was measured at 620 nm. SS was assayed as above but with 25 mm fructose instead of fructose-6-phosphate and in the absence of glucose-6-phosphate. The soluble protein content was determined by the method of Lowry[13] using bovine serum albumin as the standard. The enzyme activity was measured as micromole of sucrose or sucrose-phosphate produced per min per milligram protein.
Determination of contents of sucrose, glucose and fructose by HPLC: About 4 g of broccoli sample (for each portion) was mixed with 1 g sea sand and homogenized in a cooled mortar and pestle. Ten ml of distilled water was added to the homogenate and was centrifuged at 11 000 x g for 10 min. The mixture was filtered through a cellulose nitrate membrane filter (0.5 μm pore size). Soluble sugars were analyzed by high performance liquid chromatography (HPLC) using stainless steel column (10.7 mm IDx30 cm) packed with silica gel (gel pack C 610). The filtered water was pumped through the column at a flow rate of 1.0 ml min-1. The pressure was adjusted to 14-15 kg cm-2 and the temperature to 60°C. A RI monitor (Hitachi L-3300) was used. Sucrose, glucose and fructose were identified by their retention times and were quantified according to standards.
Statistics: A randomized complete block design was adopted with three replications. The level of significance was calculated from the F-value of ANOVA. The relationship between sugar content and enzyme activities were described by linear correlation analysis.
RESULTS
Weight loss: Percent weight loss of broccoli heads was very high after 1 d of storage in both cultivars (Fig. 1). This level rapidly decline to about 50% of the initial rate during the next 24 h. After that, no significant change was observed throughout the experimental period.
Color assessment: In both cultivars, hue angle (used as a measure of yellowing) gradually declined at the end of the storage period (Fig. 2). ‘Sairin’ showed a faster rate of hue angle decline than ‘Hartland’.
Sucrose synthase activity: In both cultivars, sucrose synthase activity gradually increased in the florets although with some fluctuations in the branchlets at the end of the storage time (Fig. 3).
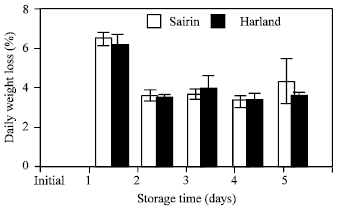 | |
| Fig. 1: | weight loss of two broccoli cultivars stored at 20°C Vertical bars indicate S.E. which, when absent, is concealed by the symbols |
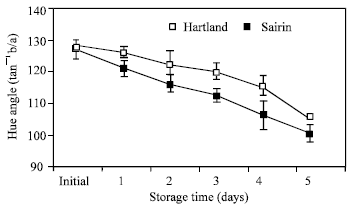 | |
| Fig. 2: | Color changes of head of two broccoli cultivars stored at 20°C. Each point represents the mean of three replications. Vertical bars indicate S.E. which, when absent, is concealed by symbols |
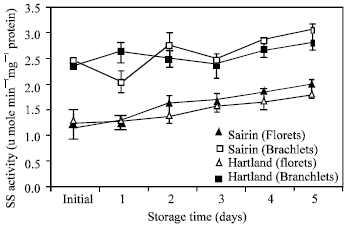 | |
| Fig. 3: | Changes in sucrose synthase activity in the florets and branchlets of broccoli stored at 20°C . Each point represents the mean of three replications. Vertical bars indicate S.E. which, when absent, is concealed by symbols |
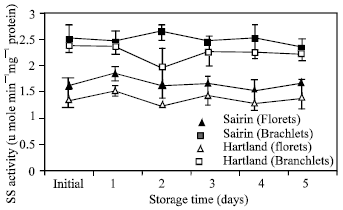 | |
| Fig. 4: | Changes in sucrose phosphate synthase activity in the florets and branchlets of broccoli stored at 20°C. Each point represent the mean of three replications. Vertical bars indicate S.E. which, when absent, is concealed by symbols |
In both cultivars, branchlets showed higher activity than florets.
Sucrose phosphate synthase activity: SPS activity did not change drastically in any portion of the two cultivars at the end of the storage period (Fig. 4). But higher SPS activity was found in the branchlets than florets.
Changes in soluble sugar content: Figure 5 shows the changes in soluble sugar content in the florets and branchlets portions of the two broccoli cultivars during storage, respectively. Sucrose content decreased gradually in the florets branchlets of two cultivars throughout the storage period while there was no remarkable change observed in glucose and fructose. A higher amount of soluble sugars (sucrose, glucose and fructose) were observed in the branchlets than florets in both cultivars. Among the three sugars, the level of fructose and glucose remained higher than that of sucrose in the florets as well as in the branchlets of the two cultivars.
Correlation coefficients (r) among enzyme activities and sugar content: Table 1 shows the correlation coefficient (r) among enzyme activities and sugar content in the florets and branchlets of two cultivars. There was a highly significant negative correlation observed between the SS activity and sucrose content in the florets and branchlets of both cultivars. There was no significant correlation between sugar contents and SPS activity in any portions of both cultivars.
DISCUSSION
Many physiological and biochemical changes occurred when broccoli is kept at ambient temperature after harvest. The major changes that we observed include color change, weight loss, sugar content and activities of sucrose metabolizing enzymes such as SS and SPS. Florets of broccoli head commenced yellowing between 3 and 4 days after storage (as indicated by decreasing hue angle values (Fig. 2). This is the most striking symptom of the postharvest senescence of broccoli which is regulated by ethylene[14]. Weight loss was highest after 1 day of storage which is due to moisture loss and loss in reducing substances of the commodity. In the present study, SS activity gradually increased in both portions of each cultivar until the end of the storage period. This increased activity may lead to degrade the sucrose level that accompanies the rapid deterioration of harvested broccoli. In many instances, it is assumed that SS is responsible for sucrose degradation[15-17].
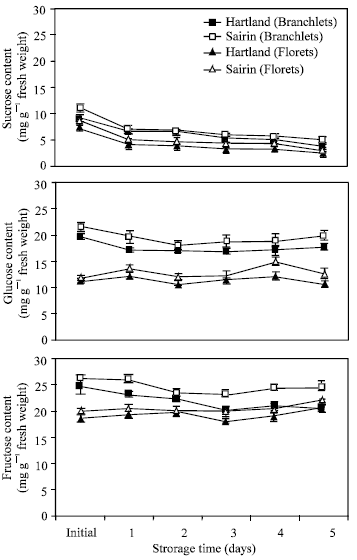 | |
| Fig. 5: | Changes in sugar content in the florets and branchlets of broccoli stored at 20°C. Each point represents the mean of three replications Vertical bars indicate S.E. which, when absent, is concealed by the symbols. |
In both cultivars, branchlets showed higher SS activity than that of the florets. Similar result is found in the bottom portion of asparagus which showed the higher SS activity than top portion[8]. The branchlets portion is composed of sink tissue. Gastonguay and Paul[18] reported that SS activity is high in sink tissue than source tissue. In branchlets soluble protein contain is lower than florets because of higher protein degradation in branchlets than florets during storage at room temperature[19-20]. On the other hand the amount of sucrose produced min was higher in florets than branchlets. This indicate that the SS activity was higher in branchlets that florets since the enzyme activity was expressed as amount of sucrose produced min-1 mg-1 of protein.
| Table 1: | Correlation coefficients (r) between the enzyme activities and sugar content in the florets and branchlets of two broccoli cultivars |
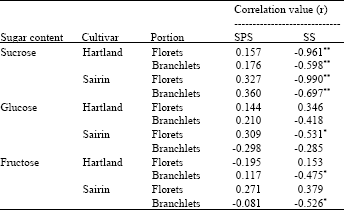 | |
| SPS= sucrose phosphate synthase, SS=sucrose synthase, *,**denote significant at p<0.05 and p<0.01, respectively, n=18 | |
There was no significant change in the SPS activity in both portions of each cultivars during storage period. Soluble sugars decline substantially throughout the storage period and concentration was higher in branchlets than florets in both cultivars. King and Morris[4] reported that soluble sugar concentration (sum of glucose, fructose and sucrose) was lower in floral section (florets) than base section (branchlets) in broccoli.
Sucrose content change rapidly but the ratio of glucose and fructose did not alter markedly in both florets and branchlets during storage period (Fig. 5). The low sucrose content could be due to an enhanced flow of sucrose to the actively respiring cell and this may cause the observed reduction in stored sucrose level seen in the broccoli.
There was a highly significant negative correlation between SS activity and sucrose content in the florets and branchlets in both cultivars (Table 1), suggesting that the decrease of sucrose content was associated with increase in the SS. Such correlation has previously been observed in asparagus spear[4]. They reported that sucrose is the major sugar which is cleaved by invertase and SS to the same direction. Based on the above study, it was found that SS cleaves sucrose in broccoli during stored at 20°C. It was also found that ‘Sairin’ and ‘Hartland’ followed similar patterns of postharvest changes in SS , SPS, sugar content and color changes but ‘Sairin’ showed higher SS, SPS activities and sugar content and became yellow earlier than ‘Hartland’.
These differences indicate that changes in SS and SPS activities play a major role in early post harvest senescence in broccoli. Further studies at the molecular level are suggested for depth understanding of the deteriorative process controlling storage sugar in broccoli during postharvest senescence.
ACKNOWLEDGMENT
The financial support of the Ministry of Education, Science, Sports and Culture of Japan under scholarship program for foreign students is gratefully acknowledged.
REFERENCES
- King, G.A. and S.C. Morris, 1994. Early compositional changes during postharvest senescence of broccoli. J. Am. Soc. Hortic. Sci., 119: 1000-1005.
Direct Link - Pramanik, B.K., T. Matsui, H. Suzuki and Y. Kosugi, 2004. Changes in acid invertase activity and sugar distribution during postharvest senescence in broccoli. Pak. J. Biol. Sci., 7: 679-684.
CrossRefDirect Link - Islam, M.S., T. Matsui and Y. Yoshida, 1996. Carbohydrate content and the activities of sucrose synthase, sucrose phosphate synthase and acid invertase in different tomato cultivars during fruit development. Sci. Hortic., 65: 125-136.
Direct Link - Bhowmik, P.K., T. Matsui, K. Kawada and H. Suzuki, 2001. Seasonal changes of asparagus spears in relation to enzyme activities and carbohydrate content. Sci. Hortic., 88: 1-9.
Direct Link - Lester, G.E., L.S. Arias and M. Gomez-Lim, 2001. Muskmelon fruit soluble acid invertase and sucrose phosphate synthase activity and polypeptide profiles during growth and maturation. J. Am. Soc. Hortic. Sci., 126: 33-36.
Direct Link - Lancaster, J.E., C.E. Lister, P.F. Reay and C.M. Triggs, 1997. Influence of pigment composition on skin color in a wide range of fruit and vegetables. J. Am. Soc. Hortic. Sci., 122: 594-598.
Direct Link - Hubbard, N.L., S.C. Huber and D.M. Pharr, 1989. Sucrose phosphate synthase and acid invertase as determinants of sucrose concentration in developing muskmelon (Cucumis melo L.) fruits. Plant Physiol., 91: 1527-1534.
PubMed - Lowry, O.H., N.J. Rosebrough, A.L. Farr and R.J. Randall, 1951. Protein measurement with the folin phenol reagent. J. Biol. Chem., 193: 265-275.
CrossRefPubMedDirect Link - Tian, M., C.G. Downs, R.E. Lill and G.A. King, 1994. A role for ethylene in the yellowing of broccoli after harvest. J. Am. Soc. Hortic. Sci., 119: 276-281.
Direct Link - Castonguay, Y. and P. Nadeau, 1998. Enzymatic control of soluble carbohydrate accumulation in cold-acclimated crowns of alfafa. Crop Sci., 38: 1183-1189.
Direct Link - Tian, MS., T. Islam, D.G. Stevenson and D.E. Irving, 1997. Color, ethylene production, respiration and compositional changes in broccoli dipped in hot water. J. Am. Soc. Hortic. Sci., 122: 112-116.
Direct Link - Page, T., G. Griffiths and V. Buchanan-Wollasten, 2001. Molecular and biochemical characterization of postharvest senescence in broccoli. Plant Physiol., 125: 718-727.
CrossRefDirect Link