
Altaf A. Dasti
Not Available
T.Z. Bokhari
Not Available
Saeed A. Malik
Not Available
Rubina Akhtar
Not Available
Asian Journal of Plant Sciences
Year: 2003 | Volume: 2 | Issue: 1 | Page No.: 42-47







ABSTRACT
The study of foliar stomata including thirty-one species belonging to three tribes of the family Boraginaceae was undertaken. The results indicated that the leaves in all the species were hypoamphistomatic and the prevalent type of stomata were anomocytic. Other types such as anisocytic, brachyparacytic, staurocytic and hemiparacytic types were occasional while paracytic, diacytic, polocytic, helicocytic, paratetracytic, actinocytic, tetracytic, brachyparatetracytic, cyclocytic and amphicyclocytic types were rare.
PDF Abstract XML References Citation
How to cite this article
Altaf A. Dasti, T.Z. Bokhari, Saeed A. Malik and Rubina Akhtar, 2003. Epidermal Morphology in Some Members of Family Boraginaceae in Baluchistan. Asian Journal of Plant Sciences, 2: 42-47.
DOI: 10.3923/ajps.2003.42.47
URL: https://scialert.net/abstract/?doi=ajps.2003.42.47
DOI: 10.3923/ajps.2003.42.47
URL: https://scialert.net/abstract/?doi=ajps.2003.42.47
Introduction
Now a days virtually every anatomical aspect of plants has been studied by taxonomists and the quantity of information accumulated is enormous. Particularly valuable taxonomic evidence has been obtained from the study of pollen, wood, leaf, epidermis, cuticle, trichomes and stomata. Some of these anatomical features are so diagnostic that they are now commonly used in routine identification, rather than being confined to a use in problems of phylogeny or classification or in the identification of fragments of plants. Trichome anatomy is of immense significance in classification at all the levels, from the circumscription of the family down to the separation of species and even varieties (Stace, 1980). Leaf epidermis and the leaf cross-sectional anatomy provide extensive taxonomic data and the literature on this subject is now vast. Characters such as the differentiation of epidermal long-cells and short-cells and the form and distribution of silica-bodies and various types of trichomes and papillae have played a big part in the modern re-classification of the family at all levels.
A number of research workers have recognized and reported unmistakable taxonomic importance of epidermal characteristics such as the shape and size of the epidermal cells, type of trichomes and type of stomata. These variations in the epidermis on the other hand, have been attributed to the functional multiplicity of the dermal tissue. The variation in the epidermal anatomy has also been studied in family Boraginaceae. Major and diverse use of stomata has been made in plant classification. Florin (1931), Metcalfe and Chalk (1950), VanCothem (1970), Dilcher (1974) and Sheikh (1980) have emphasized the systematic application of stomatal types while Shah and Gopal (1969a, b) considered the stomata as a weak point in taxonomic classification. But they feel that like other characters they can be advantageously used for that purpose. Zahur (1982) and Asghar et al. (1989) considered that the stomatal types have little taxonomic significance, because the stomatal types become modified following its development. Pant and Khare (1969) reported that the stomatal type is not constant for a taxonomic category of any rank. The reason for this, is the diversity of stomatal types in different taxa. In view of these considerations in family Boraginaceae, it was decided to undertake a survey of frequency, distribution pattern, types and percentages of open and close stomata. This family was chosen because it was found more or less in all types of climates and at all altitudes.
MATERIALS AND METHODS
Thirty-one species (15 genera, 3 tribes) belonging to the family Boraginaceae were collected from different localities of Baluchistan, i.e. WaliTangi (68.03 E x 30.13 N), Muslukh (66.77 E x 30.29 N), Hanna (67.15 E x 30.13), Quetta (67.74 E x 30.04 N), Shinghar (69.47 E x 31.43 N), (66.47 E x 31.43 N), Bibi Nani (67.40 E x 29.68 N) and Hazar Ganji (66.77 E x 30.04 N). All the species were named following the Stewart (1972). The epidermal strips of desired lengths were removed from both the abaxial and adaxial surfaces of the leaves by conventional method. These were fixed in FAA (Formaline: acetic acid: alcohol mixture) and stained with Delafield’s Hematoxyline. The stained slides were examined under the light microscope at 500x. Various anatomical features, such as number and shape of epidermal cells, number and type of stomata, length and width of guard cells, stomatal opening, types of trichomes and glands were studied. The size of the guard cells and stomatal pores were recorded with the help of a calibrated eyepiece. Ten different observations were made for each species and their mean was calculated. Other observations were recorded from the drawings, using camera Lucida adjusted at 500x microscope. The stomatal index (S.I.) was calculated using the formula adopted from Salisbury (1928) as under:
SI = X / X +Y x 100 |
| X | = | Number of stomata per unit leaf area, |
| Y | = | Number of epidermal cells per unit leaf area |
RESULTS
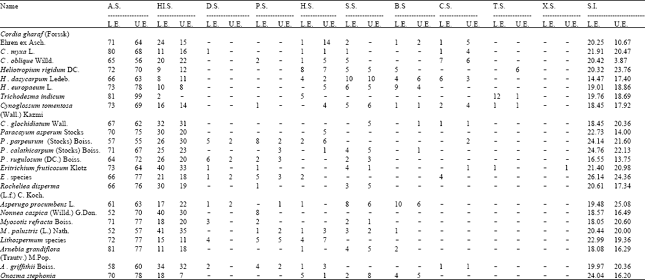
Despite the enormous diversity in the stomatal type; (with more than one type occurring even on the same surface lying side by side) the anomocytic is the dominant type of stomata (Table 1). Anisocytic type is occasional while hemiparacytic, staurocytic, brachyparacytic and cyclocytic types were quite rare.
Diacytic, paracytic, transitional from dia to paracytic, axillocytic and helicocytic types were present in the tribe Cordieae and Borageae while transitional form dia to paracytic, helicocytic and axillocytic types were present only in the tribe Borageae. Dissimilarities in the stomatal types in different species of same genera and even on the same surface of one leaf e.g. Cordia gharaf, C. myxa and C. obliqua have anomocytic, anisocytic, hemiparacytic, staurocytic and cyclocytic as the common types while diacytic and actinocytic types occur only in Cordia myxa. Paracytic and amphiparacytic types are present only in C. obliqua while C. gharaf has brachyparacytic type, which is absent in Cordia myxa and C. obliqua. Such types of diversity occur in species of other genera such as Heliotropium, Cynoglossum, Paracaryum, Eritrichium, Myosotis, Nonnea and Onosma.
Similarities of stomatal types, which encountered in species of different genera of the same tribe and even belonging to different tribes e.g. anomocytic and anisocytic types of stomata occurred in all the tribes. Cordia myxa and Arnebia grandiflora belonging to different tribes but have equal frequency of paracytic types on the lower epidermis.
All the species of the family Boraginaceae, under present study had hypoamphistomatic leaves (Table 1). The epidermal cells in the mature epidermis of all the members were irregularly arranged and were either elongated in various directions or were isodiametric in shape. The walls of epidermal cells were straight, sinuous, arched or papillose. In all the leaves, the epidermal cells on the veins or near the veins had either straight or slightly wavy cell walls. Most of the observed species had more cell wall undulations on the abaxial than adaxial ones.
All the members of the family had uniseriate common type of trichome. However, the other types were restricted to some species. Unicellular conical, calcified bristles with cystoliths present in the swollen basal portions of these hairs, were present in the species of Cordia, Trichodesma, Cynoglossum, Eritrichium, Heliotropium, Lithospermum, Myosotis and Paracaryum.
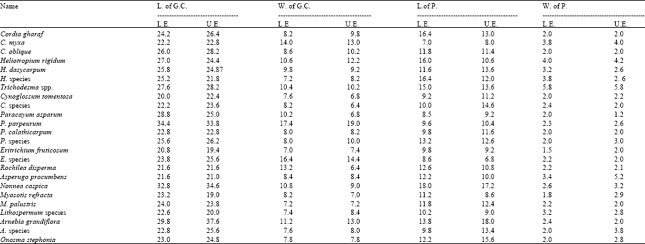
Unicellur unbranched hairs, simple, uniseriate trichomes in species of Cordia, Echium and Myosotis. Branched multicellular hairs with two equal or unequal arms were rare in species of Cordia, Nonnea and Lithospermum. Retorted shaped hairs especially on the lower surface were present in Heliotropium rigidum. Glandular hairs with short or long stalks were of various shaped, unicellular heads recorded in the species of Cordia, Echium, Heliotropium, Lithospermum, Nonnea and Onosma. Beside the stomatal types, a great diversity was observed in the size of stomata. Two fold differences were found in the width of guard cells ranging from 7.0 (Eritrichium fruticosum) to 17.4 F (Paracaryum perpeurum). Similar variations were observed in the length of guard cells ranging from 20.00 F (Cynoglossum tomentosa) to 34.4 F (Paracaryum perpeurum). Differences in the stomatal pore size were also observed between the
species (Table 2). However, all these parameters were not consistent and exhibited variations from one locality to another (Table 3).
Discussion
Results indicated that epidermal characters do not exhibit a uniform pattern and hence have very little significance as a taxonomic character within the family Boraginaceae. Interspecific differences in epidermal characters were also observed by Whang et al. (1998).
This study shows that the species studied belonging to different tribes show similar value of stomatal indices such as Cordia gharaf of the tribe Cordieae, Heliotropium dasycarpum of the tribe Heliotropieae and Paracaryum spp. of the tribe Borageae, have similar stomatal indices. Similarly different species belonging to different genera of the same tribe have similar or slightly different values of stomatal indices. Therefore, from the results of this study, it may be concluded that the stomatal index is not significant taxonomically in the species examined. Leaves of Nonnea pulla, N. nigricans and Trichodesma indicum were collected from different localities with different environmental conditions to provide comparison. Nonnea pulla collected from Kalat has higher value of stomatal index as compared to that of the specimens collected from Ziarat. It seems clear that the stomatal index is not constant and it is related to some ecological factors.
Plants collected from more than one localities show a wide variation in the stomatal types. For example the specimens of Heliotropium europaeum collected from Quetta have anomocytic, anisocytic, brachyparacytic, hemiparacytic, staurocytic, cyclocytic and actinocytic stomata as the common types. While the specimens collected from Pishin have no anisocytic but has an additional brachyparatetracytic which does not occur in the specimens collected from Quetta.
| Table 1: | Percentage of different types of stomata and stomatal index in the upper (U.E.) and lower (L.E.) epidermis |
 | |
| Table 2: | Dimension and state of stomatal complex |
 | |
| Table 3: | Spacial variations in the size of stomatal complex. All the values are the means across the ten replicates. ± indicates the standard deviation from the mean |
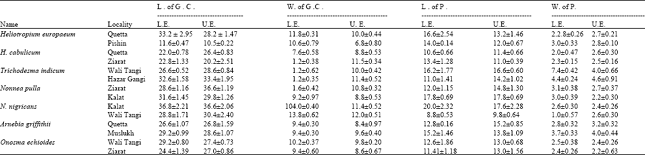 | |
In Nonnea nigricans, cyclocytic and actinocytic types were present in specimens collected from Kalat and they were absent in specimens collected from Wali Tangi. Similar modification occur in the species Heliotropium cabulicum, Trichodesma indicum, Arnebia spp., Onosma echioides, collected from different localities of diverse environment. This shows that there is no constancy of the stomatal types and modification in stomatal types occur by changing the environmental conditions.
The results of this study agree with the findings of Pant and Kidwai (1964), Inamdar (1969), Inamdar and Bhatt (1971) and Zahur (1982), who considered it a weak tool in taxonomic classification, though these can be used advantageously like other characters.
These findings go against those of Vesque (1989), Stebbins and Kush (1961), Amelunxen et al. (1967), VanCothem (1970), Kakkar and Paliwal (1972), Dilcher (1974), Raju and Rao (1977), Deghan (1980), Sen and Hannipman (1981) and Sarwar (2001) who hence regarded stomatal types as strong taxonomic character for the identification of species.
Salisbury (1928) believed that stomatal index might remain fairly constant for any one species and is a reliable indicator of the taxonomic affinity. The results disagree with his findings. The morphology of epidermal cells confirms the investigations of Metcalfe and Chalk (1950), Stace (1969) and Inamdar and Bhatt (1971). The distribution and morphology of glandular and non-glandular trichomes are likewise similar to those of Metcalfe and Chalk (1950). Trichomes are one of the most important characters for taxonomy at the level of species and genus. The studies are too numerous to list; only a few are mentioned here: Carlquist (1958), Carolin (1971), Cowan (1950), Ellis (1976), Heitzelman and Howard (1948), Metcalfe (1971), Metcalfe and Chalk (1950) and Tomlinson (1969). Even more are listed by Theobold et al. (1979).
REFERENCES
- Deghan, B., 1980. Application of epidermal morphology to taxonomic delimitations in the genus Jatropha L. (Euphorbiaceae). Bot. J. Linnean Soc., 80: 257-278.
CrossRef - Dilcher, D.L., 1974. Approaches to the identification of angiosperm leaf remains. Bot. Rev., 40: 1-157.
Direct Link - Heintzelman, C.E. and R.A. Howard, 1948. The comparative morphology of the Icacinaceae V. The pubescence crystals. Am. J. Bot., 35: 42-52.
Direct Link - Inamdar, J.A., 1969. Epidermal structure and stomatal ontogeny in some polygonales and centrospermae. Ann. Bot., 33: 541-552.
Direct Link - Inamdar, J.A. and D.C. Bhatt, 1972. Structure and development of stomata in some labiatae. Ann. Bot., 36: 335-344.
Direct Link - Kakkar, L. and G.S. Paliwal, 1972. Foliar venation and laticifers in jatropha gossypifolia. Beitrage Biol. Pflanzen, 8: 425-432.
Direct Link - Pant, D.D. and P.K. Khare, 1969. Epidermal structure and stomatal ontogeny in Eusporangiate ferns. Ann. Bot., 33: 795-805.
Direct Link - Raju, V.S. and P.N. Rao, 1977. Variation in the structure and development of foliar stomata in the Euphorbiaceae. Bot. J. Linn. Soc., 75: 69-97.
CrossRef - Salisbury, E.J., 1928. On the causes and ecological significance of stomatal frequency with special reference to the woodland flora. Phil. Trans. R. Soc. B, 216: 1-65.
Direct Link - Shah, G.L. and B.V. Gopal, 1969. Stomatal ontogeny of vegetative and floral organs of some Amaryllidaceae. Ann. Bot., 34: 737-748.
Direct Link - Stebbins, G.L. and G.S. Khush, 1961. Variation in the organization of the stomatal complex in the leaf epidermis of monocotyledons and its bearing on their phylogeny. Am. J. Bot., 48: 51-59.
Direct Link - Van Cothem, W.R.J., 1970. A classification of stomatal type. Bot. J. Linn. Soc., 63: 235-246.
CrossRef