
Muhammad Fareed Khan
Not Available
Asian Journal of Plant Sciences
Year: 2003 | Volume: 2 | Issue: 2 | Page No.: 212-215







ABSTRACT
In this protocol two primers were tested against the extracted DNA, where all the amplified genotypes were found to be similar against primer AR01. The reaction of all the genotypes under observation exhibited a series of variations against primer AR02. The plants analyzed cytologically exhibited a regular mitotic behaviour except variety sargab-92, in which 2n= 40 and 41 chromosomes have been reported. All other varieties were proved to be hexaploid with 2n= 42. High molecular weight (HMW) glutenin subunits from 7 hexaploid wheat genotypes were also fractionated by SDS-PAGE, in order to characterise the material under observation. In this experiment 6 genotypes were found to be identical, while a great variation was observed within the variety sargab-92, where some extra bands have been reported.
PDF Abstract XML References Citation
How to cite this article
Muhammad Fareed Khan, 2003. Evaluation of Hexaploid Wheat Genotypes by Using DNA Isolation and Gel-electrophoresis. Asian Journal of Plant Sciences, 2: 212-215.
DOI: 10.3923/ajps.2003.212.215
URL: https://scialert.net/abstract/?doi=ajps.2003.212.215
DOI: 10.3923/ajps.2003.212.215
URL: https://scialert.net/abstract/?doi=ajps.2003.212.215
INTRODUCTION
Isolation of plant DNA is a basic requirement for genome characterisation and also for mapping procedures involving the use of genetic markers and for the identification and isolation of plant genes for genetic engineering. The degree of purity and quality required of the DNA isolated varies from application to application. On one hand, high molecular weight, high purity DNA is required for the production of genomic DNA libraries, which are screened for plant gene sequences and for other genetic markers such as RFLPs. For genetic analysis, on the other hand, the degree of purity required may be lower, but other factors such as yield of DNA, may be more important. The lack of prior knowledge of DNA sequences and lack of characterisation of the probes restricts the use of RFLPs for detecting polymorphism.
A variety of molecular markers are now available for detecting DNA polymorphism among animal and plant materials. These markers may be a hybridization based markers or PCR based markers and include RFLPs using various types of probes: Microsatellites, Minisatellites, RAPDs and PCR (Williams et al., 1990; Welsh and Mc Clelland, 1990).
The studies of Shepherd (1968) and Wrigley and Shepherd (1973) used the aneuploids of Sears (1954) and 1- and 2-dimensional methods of gel electrophoresis to demonstrate that all gliadin protein components of hexaploid common wheat endosperm are coded by genes located on chromosomes of homeologous groups 1 and 6 (Lafiandra et al., 1984). Allelic variation of high molecular weight (HMW) subunits of glutenin in 185 cultivars of bread wheat has been described by Payne et al. (1981b), where about 20 different major subunits were distinguished by SDS-PAGE. Over the past few years, more efforts have been focused on analysis at the molecular level. Some cDNA clones have been isolated that encode LMW glutenin components (Bartels and Thompson, 1983).
The HMW subunits are closely associated with breadmaking quality (Payne et al., 1981a), but the contributions of other groups of prolamins such as gliadins and LMW glutenins are much less clear (Singh and Shepherd, 1988).
Polymorphism of glutenin subunits in 466 accessions of the wild tetraploid wheat in Israel was characterized with regard to the ecogeographical distribution of the HMW glutenin alleles, both between and within twenty two populations. (Levy and Feldman, 1987) Some populations were monomorphic for all the HMW glutenin loci, while others contained up to 4 alleles per locus. Intra-population variability could be predicted by the geographical distribution.
In this study we have compared seven Pakistani wheat varieties by using DNA isolation technique (CTAB-method) and SDS-PAGE.
MATERIALS AND METHODS
The study was conducted at the Institute of Plant Breeding and Plant Protection Martin-Luther University, Federal Republic of Germany, during a three months visit programme, sponsored by German Academic Exchange Service (DAAD). The material consisted of a collection of seven hexaploid wheat genotypes, provided by National Agricultural Research Council (NARC) Islamabad-Pakistan. The varieties comprised of: local (Rawalakot), Inqualab-91, Bakhtawar, Chakwal, Kohistan-97, Rawat-87 and Sargab-92. Plants were grown in pots inside the green house and leaf samples were harvested from seven week-old individual plants and stored in the freezer (-7oC) until DNA extraction. Each DNA preparation consisted of a single individual plant. Ten plants were used for analytical purposes. The technique followed was described by Murray and Thompson (1980).
Cytological observations: Cytological investigations were designed to check the somatic chromosome numbers and also the behaviour including the ploidy level of the indigenous wheat varieties. Preparations were made from the young seedlings grown inside the laboratory. Chromosomes were pre-treated with ice for 24 h and fixed in 3:1 (alcohol:acetic acid). The fixed root tips were hydrolysed in 1N HCl at 60 oC for 12 min, stained with fuchsin for about 15 minutes and squashed in 2% aceto-carmine.
DNA Isolation procedure (CTAB-method): This method, a modification of the procedure described by Murray and Thompson (1980) and Saghai-Maroof et al. (1984) is relatively simple and has been used successfully with a wide range of mono- and dicot plant species. This method is directly used with fresh plant material. It was generally of sufficient quality for restriction analysis or for amplification by PCR.
RNA digestion: The methodology used for DNA digestion was described previously by Saghai-Maroof et al. (1984).
DNA amplification: DNA was extracted from the young leaves using cetyl-trimethylammonium bromide (CTAB) protocol. The approximate concentration of DNA was estimated by comparison with lambda DNA (100-200 ng) in ethidium bromide stained agarose gel. Final concentration was adjusted to 20 ng μl-1 for amplification purposes.
Two primers (AR01-AR02) were tested against the extracted DNA from seven Pakistani wheat varieties. The reaction mixture was composed of 2.5 μl buffer, 2,0 μl primer, 0.50 μl dNTPs, 0.25 μl Taq polymerase and 15.75 μl water in a final volume of 21 μl. Mixture was overlayed with 25 μl oil and 4 μl DNA. The thermal cycle comprises of 6 min at 94oC, followed by 35 cycles of 1 min at 94, 1 min at 37, 2 min at 72 and a final period of extension of 5 min at 72oC followed by cooling at 4oC.
20 μl of each amplification product along with 20 μl marker was run on 1% agarose gel (200 ml 1xTAE, 2 g agarose and 10 μl ethidium bromide) for one hour and photographed under UV light.
SDS-PAGE: The variability of seed reserve protein was also analyzed in order to compare the seven Pakistani summer wheat varieties together (Khan et al., 2002).
RESULTS AND DISCUSSION
Cytological observation: The plants analyzed exhibited a regular mitotic behaviour except sargab-92, in which 2n=40 and 41 chromosomes were observed. The chromosomal count by the genotypes, local (Rawalakot), Inqualab-91, Bakhtawar, Chakwal, Kohistan-97 and Rawat-87 was 2n=42 chromosomes (Fig. 1 and 2). The plants of variety Sargab-92 shown a chromosomal variation with 2n= 40 and 41 rather than 2n=42. The similar type of alteration reported by Khan et al. (1994), who summarised the results that hexaploid wheat (Triticum aestivum L.) has a great amount of duplications and triplications of genes, due to the fact that it was formed by the addition of three closely related diploids. That is why in wheat a unique series of aneuploids is available. In order to confirm such type of results it was suggested that a meiosis study should be conducted for more critical screening of the material under observation.
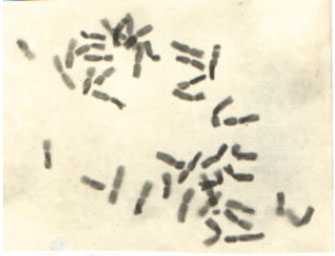 | |
| Fig. 1: | Variety Inqualab-91 with 2n= 42 chromosome |
 | |
| Fig. 2: | Metaphase I of variety Bakhtawar with 21 bivalents |
 | |
| Fig. 3: | SDS-PAGE banding patterns of 7 Pakistani wheat varieties |
| Table 1: | Comparison of amplified products (1-7) generated by primer AR01- AR02 |
 | |
| The occurrence of a specific band of amplified DNA was scored 'x' and absence as '0' for all prominent bands. | |
SDS-PAGE: As a result no variation within 6 genotypes was found and a genetic variability was observed by the genotype sargab-92, in which some extra bands were observed (Fig. 3). The diagonal arrows mark the extra bands on slot 1 of the variety sargab-92. Slot 2 and 3 seems to be similar but a clear definition is rather difficult due to the low staining intensities (Fig. 3). In order to confirm the genetic variation the whole experiment was repeated but the same results were obtained and the variation in high molecular weight (HMW) gliadin sub-unit composition of variety sargab-92 was confirmed. Allelic variation of high molecular weight subunits of glutenin in 185 cultivars of bread wheat has been observed by Payne et al. (1981b). Four hundred and sixty-six accessions of wild tetraploid wheat from Israel has been reported by Levy and Feldman (1987) in which intra-population variability is reported. Finally it was concluded that there is some thing wrong with the variety Sargab-92, which in practice needs more critical examination.
PCR: Polymerase chain action (PCR) is an in vitro method of nucleic acid synthesis by which a particular segment of DNA can be specifically replicated. The technique is an ingenious tool for molecular biology. Seven wheat genotypes were examined against 2 primers, in order to observe the difference among them. The results of an amplified DNA against 2 primers, in which a significant difference against primer AR02 was observed (Table 1). In this experiment PCR products were separated on 1% agarose gel which was stained with ethidium bromide. Hind 111 restricted lambda DNA marker was used to estimate the length of the amplified sequence.
All the amplified genotypes were found to be similar against the primer AR01, having an 800 bp fragment, while most of the varieties were shown variation against primer AR02, where local (Rawalakot) and Bakhtawar have shown a fragment length ranging from 1500-2400 bp. The variety Kohistan-97 has only one band with 2400 bp fragment length. All other varieties exhibited an equal amount of response towards AR02 having 2000-2400 bp fragment length. Similar type of results were obtained by Welsh et al. (1991). He used four colonies for PCR amplification purposes. Out of 4 recombinants colonies used 3 (1, 2 and 4) showed insert sequence length of 1664, 1733 and 1800 bp respectively. Third colony showed a slight less insert size (1400 bp).
During this course of investigations all the objectives could not be achieved due to the short period of stay at the Martin-Luther- University, Federal Republic of Germany, therefore, further investigations are recommended for confirmation of the results.
ACKNOWLEDGMENT
Thanks are due to German Academic Exchange Service (DAAD) for financing this short visit programme to Federal Republic of Germany. I am also thankful to Prof. Weber, who graciously provided me the laboratory facilities during this course of investigations.
REFERENCES
- Bartels, D. and R.D. Thompson, 1983. The characterisation of cDNA clones coding for storage proteins. Nucleic Acid Res., 11: 2961-2971.
Direct Link - Levy, A.A. and M. Feldman, 1988. Ecogeographical distribution of HMW-glutenin alleles in populations of the wild tetraploid wheat Triticum turgidum var. dicoccoides. Theor. Applied Genet., 75: 651-658.
CrossRefDirect Link - Murray, M.G. and W.F. Thompson, 1980. Rapid isolation of high molecular weight plant DNA. Nucl. Acids Res., 8: 4321-4326.
CrossRefPubMedDirect Link - Payne, P.I., K.G. Corfield, L.M. Holt and J.A. Blackman, 1981. Correlations between the inheritance of certain high-molecular-weight subunits of glutenin and breadmaking quality in progenies of six crosses of bread wheat. J. Sci. Food Agric., 32: 51-60.
CrossRef - Payne, P.I., L.M. Holt and C.N. Law, 1981. Structural and genetical studies on the high-molecular-weight subunits of wheat glutenin. Part I: Alleic variation in subunits amongst varieties of wheat (T. aestivum). Theor. Applied Genet., 60: 229-236.
CrossRefDirect Link - Saghai-Maroof, M.A., K.M. Soliman, R.A. Jorgensen and R.W. Allard, 1984. Ribosomal DNA spacer-length polymorphisms in barly: Mendelian inheritance, chromosomal location, and population dynamics. Proc. Natl. Acad. Sci., 81: 8014-8018.
CrossRefDirect Link - Singh, N.K. and K.W. Shepherd, 1988. Linkage mapping of genes controlling endosperm storage proteins in wheat. Theor. Applied Genet., 75: 642-650.
CrossRef - Welsh, J., R.J. Honeycut, M. McClelland and B.W.S. Sorbral, 1991. Parentage determination in maize hybrids using the arbitrarily primed polymerase chain reaction (AP-PCR). Theoret. Applied Genet., 82: 473-476.
CrossRefDirect Link - Welsh, J. and M. McClelland, 1991. Genomic fingerprinting using arbitrarily primed PCR and a matrix of pairwise combinations of primers. Nucleic Acids Res., 19: 5275-5279.
Direct Link - Williams, J.G.K., A.R. Kubelik, K.J. Livak, J.A. Rafalski and S.V. Tingey, 1990. DNA polymorphisms amplified by arbitrary primers are useful as genetic markers. Nucleic Acids Res., 18: 6531-6535.
CrossRefPubMedDirect Link - Wrigley, C.W. and K.W. Shepherd, 1973. Electrofocusing of grain proteins from wheat genotypes. Ann. N. Y. Acad. Sci., 209: 154-162.
PubMedDirect Link