
Abd El-baky
Not Available
H. Hanaa
Not Available
Mohamed, A. Amal
Not Available
M.M. Hussein
Not Available
Asian Journal of Plant Sciences
Year: 2003 | Volume: 2 | Issue: 17 | Page No.: 1220-1227







ABSTRACT
The changes that take place in the leaf apoplastic antioxidant enzymes system, protein and isoenzyme pattern in response to salts stress was studied in three onion (Allium ceba L.) cultivars, named Behary Red, Giza 6 and Giza 20. The constitutive level of lipid peroxidation and glutathione content (GSH), enzymes activity of catalase (CAT), superoxide dismutase (SOD) and peroxidase (POD) as well as the protein and isoenzymes pattern were performed in the leaf of control (tap water, 200 ppm salts) and three concentrations of salt (2000, 4000 and 6000 ppm salts prepared from seawater (28000 ppm salts) stressed plants, 45 days after completing the stress treatments. The salt stress increased the rates of lipid peroxidation (as indicated by increasing the malonaldehyde (MDA) contents) and their effect was increased gradually by increasing salts levels. The increasing values in MDA contents in salt stress plants treated at 2000, 4000 and 6000 ppm salts levels were 121.9, 248.6 and 290.3% in Behary Red cv. and 126.4, 260.0 and 396.2% in Giza 20 cv., respectively of the control level (100%). Generally, salts stress induced significant increase in the activities of the antioxidative enzymes CAT, POD and SOD in onion cvs., accompanied by increased in glutathione level. The GSH level was increased more in Behary Red cv. and Giza 20 cv., than in Giza 6 cv., at 6000 ppm salts. Their increasing values were 203.7, 283.3 and 242.86%, respectively compared to the control level (100%). The leaf protein profile obtained by SDS-PAGE was diagnostic of characteristics different onion cultivars exposed to salts stress which, three distinct new protein bands with molecular weigh 50, 66 and 80 kDa were produced in Giza 6, Giza 20 and Behary Red onion cultivars, respectively. Also, electrophoretic bands of POD and CAT isoenzyme of different onion cultiver were differ in numbers and relative concentration due to salts stress. This finding suggested that Behary onion cultiver is better protected against lipid peroxidation process, inherently and under salts stress, than Giza 6 and Giza 20 onion cultivers. Also, the new protein bands and the characteristic CAT and POD isoenzymes banding patterns could be used as a biochemical marker for selection of salt tolerance onion plants when grown under salts stress.
PDF Abstract XML References Citation
How to cite this article
Abd El-baky, H. Hanaa, Mohamed, A. Amal and M.M. Hussein, 2003. Influence of Salinity on Lipid Peroxidation, Antioxidant Enzymes and Electrophoretic Patterns of Protein and Isoenzymes in Leaves of Some Onion Cultivars. Asian Journal of Plant Sciences, 2: 1220-1227.
DOI: 10.3923/ajps.2003.1220.1227
URL: https://scialert.net/abstract/?doi=ajps.2003.1220.1227
DOI: 10.3923/ajps.2003.1220.1227
URL: https://scialert.net/abstract/?doi=ajps.2003.1220.1227
INTRODUCTION
Recently, the environmental stresses such as drought, temperature, air pollution, heavy metals, pesticides and soil pH (Prasad et al., 1994; Zhang and Kirkham, 1996; Badiani et al., 1993; Gossett et al., 1996) are major factors limiting crop production due to its affects almost all plant functions (Hern-Ndez, et al., 2001). Salt stress causes stomatal closure, which reduces the CO2 to O2 ratio in leaves and inhibits CO2 fixation, these conditions increase the rate of reactive oxygen species (ROS) formation, via enhanced leakage of electron to oxygen in the chloroplasts and mitochondria. Electrons generated by photosynthetic process react with oxygen and generate ROS● such as superoxide (O2¯) peroxide (H2O2), hydroxyl radicals (●OH) and singlet oxygen (1O2) (Asada and Takahashi, 1987). Also, the superoxide radical is at the membrane level in most plant cell organelles and hydrogen peroxide is the product of superoxide dismutase of several oxidases of peroxisomes (Thompson et al., 1987; Del-Rio et al., 1992; Shalata and Tal, 1998). The hydroxyl radical can be generated through the Fe-catalyzed Haber-Werss reaction (Halliwell and Gutteridge 1985) and singlet oxygen is mainly formed in photodynamic process in the chloroplast (Thompson et al., 1987). However, during normal condition, ROS● are efficiently scavenged by antioxidant system. While, when ROS? production is surpasses the antioxidant system capacity, oxidation stress occur, resulting in cytotoxic protein damage, DNA damage and lipid peroxidation (Elstner 1982; Shalata and Tal, 1998).
Plants have evolved a battery of antioxidative mechanisms to detoxify and eliminate these reactive oxygen species. The antioxidant defense system includes hydrophilic (ascorbic acid and glutathione), hydrophobic (α-tocopherol and carotenoids) antioxidants substance and enzymes like superoxide dismutase, catalase, peroxidase and others. Plants differ in their ability to remove and scavenge ROS●. The differences in plant protection mechanisms determine their tolerance to stress conditions associated with ROS toxicity. Salt stress caused a reduction in CAT activity in some plants, e.g., pea (Corpas et al., 1993) and rice seedling (Mittal and Dubey, 1992) while, in other plants, salt stress enhanced CAT activity e.g., in seedling of Cicer arietinum (El-Kady et al., 1982). Salt induced a significant increase in a SOD activity in citrus and pea (Gueta-Dahan et al., 1997). However, salt-induced increase of antioxidant enzymes which is presumed to be active in elimination of hydroperoxidants, either constitutive or induced have been experimentally determined to exhibit greater tolerance to variety of environmental stress conditions (Lechno et al., 1997; Shalata and Tal, 1998).
When onion is grown for bulbs it is classified as low salt tolerant crop (Mangal et al., 1989). In this study the antioxidant enzyme activity (SOD, CAT and POD) as well as the glutathione (GSH) and lipid peroxidation contents were studied in the young leaves of onion cultivars (Giza 6, Giza 20 and Behary Red) when grown under salt stress condition in order to select salt tolerant cultivar.
MATERIALS AND METHODS
Plant materials: Three onion cultivars named: Behary Red, Giza 6 and Giza 20 were obtained from Onion Department, Agriculture Research Center, Giza, Egypt.
Growth conditions: A pot experiment was conducted in the greenhouse of National Research Center, Dokki, Cairo, Egypt during 2001/2002-winter season in order to evaluate the effect of different salt concentration (prepared by diluting seawater) on the lipid peroxidation, antioxidant enzyme activities, protein pattern and isoenzyme profiles as follows:
Salt concentration: Tap waters (200 ppm salts) used as control and diluting seawater (2000, 4000 and 6000 ppm salts), the experiment included 12 treatment and the experimental design was split plot in 10 replicated. Seedlings of the onion cultivars were transplanted after 40 days from sowing. Salt stress treatments were started at 60 days from sowing which plants irrigated by saline water and in next irrigation followed by tap water alternatively. Samples from every treatment were taken at 45 days from transplanting for chemical analysis. At the same time the plant received the optimum fertilizer (PO-4, K+ and N).
Preparation of cytosolic fraction: Leaves tissue of onion cultivars about (1 g) were excised and homogenized using mortar and pestle in 4 ml of ice-cold grinding buffer containing 250 mM sucrose, 25 mM Tris and the pH was adjusted to 7.2. The homogenate was centrifuged at 12000 g for 15 min at 4°C. The resulting supernatant was used for analyses of enzyme activities, GSH, lipid peroxidation, protein content and isoenzymes electrophoresis patterns.
Enzymes assays: The activity of cytosolic SOD (EC 1.15.1.1) was determined by photochemical method (Ginnopolitis and Ries, 1977). Spectrophotometrically method as described by Chance and Maehly (1955) was used to assay the POD activity (EC 1.11.1.7). The CAT enzyme (EC 1.11.1.6) activity was measured spectrophotometrically by measuring the U.V absorption at 240 nm of a 10 mM of H2O2 solution in present of 25 mM phosphate buffer, pH 7.0 (Hans- Luck 1970).
Determination of lipid peroxidation products: The lipid peroxidation products was estimated by the formation of thiobarbaturic acid reactive substances (TBARS) and quantified in term of malonaldheyde (MDA) as described by Haraguchi et al. (1997). The lipid peroxidation was expressed as micromoles of MDA. The extinction coefficient of TBARS was taken as 1.56 x 10 5 mM at wave length 532 nm.
Determination of glutathione (GSH): The GSH content was estimated by the acid–soluble sulfhydryl (SH) level in the tissue homogenates as described by Vecchia et al. (1992).
Determination of protein: Protein levels of cytosolic were determined spectrophotometrically at 595 nm, using comassie blue G 250 as a protein binding dye (Bradford, 1976). Bovine serum albumin (BSA) was used as a protein standard.
Protein electrophoresis: In order to determine the molecular weight of the protein subsets, sodium dodycyl sulphate polyacrylamide gel electrophoresis (SDS-PAGE) under denaturating conditions were used as described by Laemmli (1970) using 12.5% acrylamide concentration. Each sample (protein extract) was heated at 100°C for 2 min in 10 mM Tris HCl buffer, pH 7 that containing 2% (w/v) SDS, 2% 2-mercapoethanol and 5% (W/V) glycerol, before electrophoresis. A 30 μl of the prepared sample were applied per well after adding one drop of the loading buffer bromophenol blue and glycerine (Gel was run at 200 volt under cooling) when the samples reach to the end of gel, the gels were removed from the apparatus, then immersed in staining solution containing comasie brilliant blue-R250. Molecular weight marker SDS-7 (Sigma) was used to estimate the molecular weight of the sample ranging from 14-66 KD.
Detection of isozyme by electrophoresis: Native-PAGE Non-denaturing polyacryiomide gel electrophoreses was done in 7.5% polyacrylamide according to Davis (1964) on vertical slabs gel using the pharmica gel electrophoresis model at 4°C using 200 volt for 120 min. Protein extracts (20 μg protein equivalent) of each sample were loaded into each well at the top of gel. Activity stain for each enzymes was carried out as follows:
POD: Gel was stained for POD activity as described by Schrauwen (1966). Gel was incubated in 0.3 mM Na-phosphate buffer, pH 6 for 10 min. Then the gel were treated with 0.1% hydrogen peroxide and 0.25% O-dianisidine.
The appearance of dark brown bands was caused by peroxidase activity and indicated the presence of a peroxidase isozymes in the gel. Then stopped the reaction with tap water and the gel photogeraphed immediately.
CAT: CAT was detected by method of Woodbury et al. (1971), the gel was soaked in 5 mM hydrogen peroxide solution. After 10 min the gel was washed with water and stained with a reaction mixture containing 2% (w/v) ferricyanide and 2% ferric chloride. The enzyme appeared as yellow bands on dark green back ground. The reaction was stopped with tap water and the gel were photographed immediately.
RESULTS AND DISCUSSIONS
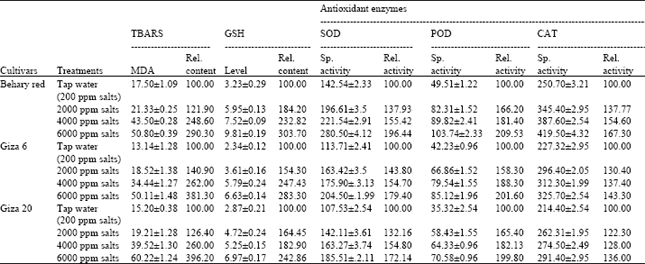
Salt stress and lipid peroxidation: The lipid peroxidation product (as MDA) was lower in Giza 6 than Giza 20 and Behary Red cvs. Under control condition, which their values were 13.1,15.2 and 17.5 μM, respectively. Their values increased in their cultivars when grown under salt stress (Table 1). The increasing values of MDA in Behary Red cv. was lower than the other two cultivars at all salt stress treatments, the increasing level of TBARS at 2000, 4000 and 6000 ppm salts were 121.9,248.6 and 290.3% in Behary Red ; 140.9,262 and 381.3% in Giza 6 and 126,260 and 396.2% in Giza 20, respectively compare to the controls level (100%). However, salt stress at highest level of 4000 and 6000 ppm salts caused a higher significant increase in levels of MDA content in all cultivars. The degree of accumulation of MDA in onion leaves was arranged in follow ascending order: Behary Red, Giza 20 and Giza 6 cv. consequently, Behary Red cv. was more salt tolerance than the Giza 20 and Giza 6 cultivars, which, the accumulation of MDA was more in the salt susceptible than in the salt tolerant cultivars (Sreenivasulu et al., 1999). Thus, the salt stress caused an increase the rate of lipid peroxidation in the susceptible cultivars (Giza 20 and Giza 6 cultivars).
Salt stress and antioxidant enzyme activities: Activities of the antioxidive enzymes CAT, POD and SOD were enhanced by salts treatment of all onion cultivars plants (Table 1). Behary Red cultivar had the higher SOD, CAT and POD activity than the other onion cultivars grown under salt stress. The values were higher in Behary Red cv. than in other two cultivars under control condition (200ppm salts). The increasing% of SOD, POD and CAT activity in Behary Red cv. grown under 2000,4000 and 6000 ppm salts were 137.93, 166.2 and 137.77%, 155.42, 181.4 and 154.6% and 196.44, 209.53 and 167.3% respectively, of the control values (100%).
The increasing % of SOD, POD and CAT activities which were lower in the Giza 20 cv. than Behary Red and Giza 6 cv. under salt stress condition. For instance, the increasing % of CAT activity grown in 2000, 4000 and 6000 ppm salts was 122,128,136% while, in Behary Red was 137,154 and 167% respectively, of the control level 100%. However, salt stress was enhancement the activity of CAT in all onion cv. were over than other enzymes SOD and POD. The means increase values of their enzymes in Behary Red cv. and Giza 6 in parenthesis grown under salt stress were 350.8, (26.7), 210.3 (149.6) and 81.3 (57.14) respectively. Where, mean values activity of POD, SOD and CAT in Giza 6 cv. was 290.4,164.6 and 68.44, respectively.
The qualitative and quantitative changes in the activity of several enzyme including SOD, CAT, POD and glutathione reductase (GR) and peroxidase (GPX) isolated from plants subjected to salinity stress were reported by Sreenivasulu et al. (1999).
Antioxidant content of glutathione (GSH) in different onion cv. treated with salts stress was shown in Table 1. Glutathione content was higher in leaves of different onion cv. grown under salt stress. The increased of GSH content was gradually increased by increasing of salts levels. At 6000 ppm salts, the level of GSH was increased by 303.7 283.3 and 242.8% of the control 100% in Behary Red cv., Giza 6 and Giza-20 onion cvs., respectively. However, GSH may play a protective role in scavenging of single oxygen, peroxides and hydroxyl radicals and is involved in recycling reduced of ascorbic acid (ASC) in the ascorbate-glutathione pathway in chloroplasts (Foyer, 1993).
| Table 1: | Effect of salt stress on lipid peroxidation (TBARS) ,Glutathione (GSH) , superoxid dimutase (SOD), peroxidase ( POD) and catalase (CAT.) of different onion cultivars |
 | |
The increasing content of GSH may be driven by enhanced of H2O2 formation in the salt stress (Lechno et al., 1997). The increasing of H2O2 level has been shown to enhance antioxidant content and antioxidative enzyme in many plants (Mitler and Tel-Or, 1991 and Lechno et al., 1997).
These results revealed that Behary Red cv. had much lower of lipid peroxidation product (MDA) and higher antioxidant (GSH) content and antioxidant enzymes activity, than the other onion cvs.. GSH content and antioxidant enzyme activities were reporting to play a key role in salt tolerance (Shalata and Tal, 1998). A regulated balance between oxygen radical production and destruction required, if metabolic efficiency and function are to be maintained ether in normal or stress condition. A constitutively high antioxidant capacity under stress conditions with plant resistance to that particular stress. Consequently, the mechanisms that reduce ROS● species and increase antioxidant enzyme system in plant are an important role in imparting tolerance in plant under environmental stress (Sreenivaslu et al., 1999 and Lechno et al., 1997).
In the present study, a significant elevation in antioxidant enzymes activities (SOD, POD, CAT) and antioxidant content (GSH) were recorded in three onion cvs. under salts stress condition. Further the degree increases were found to be correlate with salts levels (Fig. 1). Furthermore, the degree of elevation in antioxidant enzymes activity and antioxidant content were relative high in Behary Red cv. than that in Giza 20 and Giza 6 cvs., therefore, Behary Red cv. can be considered as better tolerant cultivars when compared to other both cultivars.
The proportional contribution of various companions of the antioxidative system in plant under salt stress may vary in different species. In cotton the results suggest that protection for oxidation damage by higher level of antioxidant enzyme like CAT and POD more active ASC-GSH care involved in development of salt stress (Gossett et al., 1996). While, the activity of SOD, inherited or induced was fond to be higher in salt-tolerant cultivars of pea or citrus than the sensitive cultivars (Gueta-Dahan, 1997). An increase in POD activity is common response to various oxidative stress factors. The tolerant cultivar (cv Prasad) of fox-tail millet had low MDA content and high POD activity than the susceptible cultivars during salinity stress.
In conclusion, salts stress onion plants exhibited specific responses of enhanced CAT, POD and SOD activities, accompanied by increase in GSH content and decrease lipid peroxidation process and their responses vary between cultivars.
Protein SDS-PAGE patterns: The protein patterns of three onion cultivars which grown at different salt treatments were analyzed. The separated bands of protein subunits were photographed and presented in Fig. 2. In general, all cultivars under all treatments contained one major distinct band with high molecular weight 66 KD. The pattern of high molecular weight band indicated that, there was a particular strong induction of biosynthesis of 66 KD protein in all cultivars under different salt treatments. In addition, in case of the first cultivar (Behary Red) which presented in lanes 1, 2, 3 and 4 respectively, produced similar number of bands except in lane 1(control treatment) which had bands differ in their intensities.
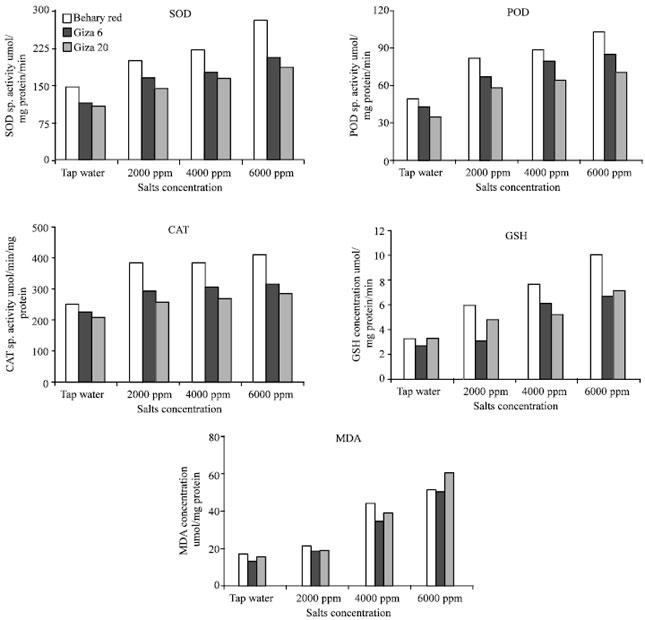 | |
| Fig. 1: | Effect of salt stress on superoxid dimutase (SOD), peroxidase ( POD), catalase (CAT), Glutathione (GSH) and lipid peroxidation(MDA) of different onion cultivars |
In contrast, in case of the second cultivar (Giza 6) different patterns of protein profile were observed, for example in lane 8 with 6000 ppm salts treatment there are two distinct new bands with MW 50 KD and 14 KD which not exhibited in any other culivars. In general, these patterns may give remarkable marker to relay the discrimination on between treated and untreated cultivar since one could considered the presence of new bands as an adaptive band for stress treatment.. Also the third cultivar (Giza 20) exhibited a new faint band with MW≅80 KD (lane 10) under 2000 ppm salt stress treatment. Consequently these newly synthesized bands indicated that salt stress induced related gene(s) to produces these salt inducible proteins (Abdel-Tawab and Allam 2002). These results are in agreement with the findings of Ericson and Alfinito (1984) who stated that, two different protein bands with MW of 32 and 20 KD were exhibited with high intensity in the salt stressed tobacco plants. El-Farash et al. (1993) studied the expression of 12 different proteins bands, which were induced in salt stressed tomato plants. They reported that the expression of these proteins bands was genetically regulated, depending on the salt concentration.
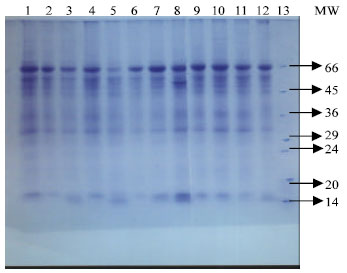 | |
| Fig. 2: | SDS-PAGE pattern of 3 different onion cultivars |
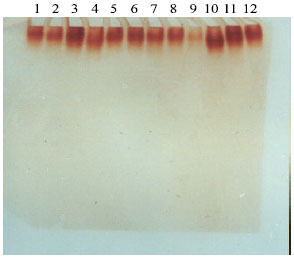 | |
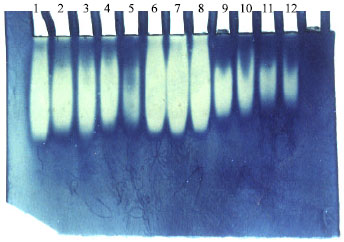
| Fig. 3: | Zymogram of POD Isoeyme of 3 different onion cultivars |
Lanes 1 to 12 where:
Behary Red
1-Control (tap water 200 ppm salts)
2-2000 ppm salts
3-4000 ppm salts
4- 6000 ppm salts
Giza 6
5-Control (tap water 200 ppm salts)
6-2000 ppm salts
7-4000 ppm salts
8- 6000 ppm salts
Giza 20
9-Control (tap water 200 ppm salts)
10-2000 ppm salts
11-4000 ppm salts
12- 6000 ppm salts
Peroxidase (POD) isozyme expression under salt stress: Expression of the POD isoenzyme was detected in leaf water soluble proteins extracted from three onion cultivars treated with 200, 2000, 4000, 6000 ppm salts using 7.5% native-PAGE system (Fig. 3).
 | |
| Fig. 4: | Zymogram of CAT Isoeyme of 3 different onion cultivars |
The results showed that one band were exhibited in salt untreated and treated plants of all cultivars. This band was manifested higher densities and intensities in the salt treated cultivars than in cultivars grown under control conditions (200 ppm salts).In the salt treated Giza 20 cultivras, the density and intensity of this band was higher under 2000, 4000, 6000 ppm salts treatments (lanes 10, 11 and 12 respectively compared to the control (200 ppm salts). These results indicated that salt stress increased the accumulation of the POD enzyme and that encoding gene(s) which may accelerated in response to salt stress in both treated and untreated treatments, but the rate of acceleration was markedly higher in the former than in the latter case. These results are in harmony with the findings of Sreenivasulu et al. (1999) who stated that high peroxidase isozyme activity was detected in salt tolerant cultivar compared to salt susceptible cultivar of Fox-tail millet which related to the salt adaptation process.
Catalase (CAT) isozyme expression under salt stress: The electrophoretic patterns of catalase isozymes was photographed and illustrated in Fig. 4. Looking at all cultivars when grown under control and salt stress treatments, each cultivar exhibited one band as a total number with different intensity. The first cultivar exhibited one band with high intensities when grow under control treatment (Lane 1) and the intensity was decreased with increasing salt treatment but still higher than any other treatment. In this concern El-Baz et al. (2003) used peroxidase isozyme and protein pattern as a marker for salts stress tolerance in cucumber plants, the authors found that, the profile of isoperoxidase was modified during salt stress conditions also a new subset of proteins induced by salt stress compared to control plant was observed. This behavior may be due to its ability to tolerate salt stress or due to the effect of salt stress which may cause some shift in gene expression. It is also interesting to note that the last cultivar showed differences in the mobility of bands and its intensity under salt treatment when compared with control.
In conclusion the present study demonstrated that, the biochemical and molecular markers associated well with salinity stress in onion cultivars. It offers a simple and fast technique could be used to select the salt tolerant culivars of onion plant.
REFERENCES
- Badiani, M., G. Schenone, A.R. Paolacci and I. Fumagalli, 1993. Daily fluctuations of antioxidants in been (Phaseolus vulgaris L.) leaves as affected by the presence of ambient air pollutants. Plant Cell Physoil., 34: 271-279.
Direct Link - Del-Rio, L.A., L.M. Saudalio, J.M. Palma, P. Bueno and F.J. Corpas, 1992. Metabolism of oxygen radicals in peroxisomes and cellular implications. Free Radical Med. Biol., 13: 557-580.
Direct Link - El-Baz, F.K., A.A. Mohamed and A.A. Aly, 2003. Development of biochemical markers for salt stress tolerance in cucumber plants. Pak. J. Biol. Sci., 6: 16-22.
CrossRefDirect Link - Elstner, E.F., 1982. Oxygen activation and oxygen toxicity. Ann. Rev. Plant Physiol., 33: 73-96.
Direct Link - Ericson, M.C. and S.H. Alfinito, 1984. Proteins produced during salt stress in tobacco cell culture. Plant Physiol., 74: 506-509.
Direct Link - Gueta-Dahan, Y., Z. Yaniv, B.A. Zlinskas and G. Ben-Hayyim, 1997. Salt and oxidative stress: Similar and specific response and their relation to salt tolerance in citrus. Planta, 203: 260-469.
PubMedDirect Link - Haraguchi, H., H. Ishikawa and I. Kubo, 1997. Antioxidative action of diterpenoids from Podocarpus nagi. Planta Med., 63: 213-215.
CrossRefPubMedDirect Link - Hernandez, J.A., M.A. Ferrer, A. Jimenez, A.R. Barcelo and F. Sevilla, 2001. Antioxidant system and O2/H2O2 production in the apoplast of pea leaves: Its relation with NaCl-induced necrotic lesions in minor veins. Plant Physiol., 127: 817-831.
Direct Link - Laemmli, U.K., 1970. Cleavage of structural proteins during the assembly of the head of bacteriophage T4. Nature, 227: 680-685.
CrossRefDirect Link - Lechno, S., E. Zamski and E. Tel-Or, 1997. Salt stress-induced responses in cucumber plants. J. Plant Physiol., 150: 206-210.
Direct Link - Prasad, T., M. Anderson, B.M. Martin and C.R. Stzward, 1994. Evidence for chilling-induced oxidative stress in maize seedling and regulatory role for hydrogen peroxide. Plant Cell, 6: 65-74.
Direct Link - Shalata, A. and M. Tal, 1998. The effect of salt stress on lipid peroxidation and antioxidants in the leaf of the cultivated tomato and its wild salt-tolerant relative Lycopersicon pennellii. Physiologia Plantarum, 104: 169-174.
CrossRefDirect Link - La Vecchia, C., E. Negri, S. Franceschi, B. D'Avanzo and P. Boyle, 1992. Tea consumption and cancer risk. Nutr. Cancer, 17: 27-31.
CrossRefPubMedDirect Link - Woodbury, W., A.K. Spencer and M.A. Stahmann, 1971. An improved procedure using ferricyanide for detecting catalase isozymes. Anal. Biochem., 44: 301-305.
CrossRefDirect Link - Zhang, J. and M.B. Kirkham, 1996. Lipid peroxidation in sorghum and sunflower seedlings as affected by ascorbic acid, benzoic acid and propyl gallate. J. Plant Physiol., 149: 489-493.
CrossRefDirect Link - Giannopolitis, C.N. and S.K. Ries, 1977. Superoxide dismutases. I. Occurrence in higher plants. Plant Physiol., 59: 309-314.
CrossRefPubMedDirect Link - Davis, B.J., 1964. Disc electrophoresis. II. Method and application to human serum proteins. Ann. N. Y. Acad. Sci., 121: 404-427.
Direct Link - Bradford, M.M., 1976. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem., 72: 248-254.
CrossRefPubMedDirect Link