
T. Ramasubramanian
Not Available
A. Regupathy
Not Available
Asian Journal of Plant Sciences
Year: 2003 | Volume: 2 | Issue: 17 | Page No.: 1170-1174







ABSTRACT
The relative fitness of pyrethroid selected populations of Helicoverpa armigera was studied by construction of age specific fecundity tables for F2,F7 and F14 generations of unselected, fenvalerate and cypermethrin selected populations. The net reproductive rate computed from the life table statistics indicated that the fitness of the pyrethroid selected populations decreased drastically as the population advanced to fourteenth generation. In F2 the net reproductive rate was 237.437, 231.359 and 228.24 for the unselected, fenvalerate and cypermethrin selected populations respectively. It came down to 123.677 and 121.432 for fenvalerate and cypermethrin selections respectively and only a marginal decline to 197.962 in unselected population. The decrease in fitness was greatly manifested through reproductive behaviour but not by the developmental traits. The variation in generation time was insignificant between the selected and unselected populations irrespective of the generations.
PDF Abstract XML References Citation
How to cite this article
T. Ramasubramanian and A. Regupathy, 2003. Relative Fitness of Pyrethroid Selected Populations of Helicoverpa armigera Hub. Asian Journal of Plant Sciences, 2: 1170-1174.
DOI: 10.3923/ajps.2003.1170.1174
URL: https://scialert.net/abstract/?doi=ajps.2003.1170.1174
DOI: 10.3923/ajps.2003.1170.1174
URL: https://scialert.net/abstract/?doi=ajps.2003.1170.1174
INTRODUCTION
The cotton bollworm, Helicoverpa armigera (Hubner) is considered as one of the major threats to the present day intensive agriculture in India and other countries in view of its direct impact on the yields of many field crops of varied nature. There are 60 cultivated species serve as host plants for H.armigera (Reed and Pawar, 1981). Control of this pest is generally achieved with insecticides more specifically with synthetic pyrethroids. More than 75% of the insecticides used in cotton are being targeted towards H.armigera (Banerjee et al., 2000), of which synthetic pyrethroids alone constitute 50-70% (Jayaswal, 1989). This Indiscriminate use of pyrethroids led to the development of resistance in H.armigera. At present, the resistance to pyrethroids is ubiquitous across the Indian subcontinent (Armes et al., 1996). Reports on the relative fitness of pyrethroid resistant population of H.armigera are however very scarce. Hence, a research was carried out to study the impact of pyrethroid selection on biological and reproductive traits of H.armigera.
MATERIALS AND METHODS
Mass rearing of H.armigera: H.armigera larvae collected from fields were pooled and divided into two sets. One set was allotted for exposure to pyrethroids and other for non-exposure. In case of exposure to pyrethroids, the surviving larvae treated with pyrethroids were reared to next generation. In case of non-exposure, unexposed larvae were reared through next generation. Both the pyrethroid resistant and susceptible populations were reared continuously on chickpea based semi synthetic diet (Armes et al., 1993) over fourteen generations.
Developing resistant strains: The selection pressure was created by two different methods.
Discriminating doses of representative pyrethroids (Fenvalerate- 0.2 μg; Cypermethrin 0.1 μg) were applied topically to create selection pressure in the first generation and the doses were gradually increased @ 0.1 μg for each generation up to third.
The LD50 of F3 unselected population (3.231 μg for fenvalerate and 3.249 μg for cypermethrin) were subsequently used to create selection pressure in the respective resistant populations from F4 onwards. The resistant strains of fenvalerate and cypermethrin were developed by retaining the survivors of the respective synthetic pyrethroids.
Construction of Age specific fecundity tables: The relative fitness of pyrethroid selected strains was studied by construction of age specific fecundity tables for F2, F7 and F14 generations as per the standard format of Birch (1948) using computerized programme. Several population growth indices viz., intrinsic rate of increase, finite rate of increase, net reproductive rate, weekly multiplication rate, mean generation time and doubling time were estimated for both pyrethroid selected and unselected strains of H. armigera using the following formulae:
| 1. | Net reproductive rate (R0) = ∑ lx mx |
| 2. | Mean generation time |
 |
| 3. | The capacity for increase |
| 4. | Intrinsic rate of increase or innate capacity of increase or infinitesimal rate of increase (rm): The rc is an approximate value only. A more accurate value of intrinsic rate of increase rm was calculated by the Graphical method suggested by Southwood (1966) using computerized programme. From the rm value, the corrected generation time (T), finite rate of increase (λ) and weekly multiplication rate (WMR) were derived by using the following formulae. |
| 5. | Corrected generation time (T) = Loge Ro / rm |
| 6. | Finite rate of increase (λ) = anti Loge rm. |
| 7. | Weekly rate of multiplication = (erm)7. |
| 8. | Doubling time (DT) in days = loge 2/rm |
RESULTS AND DISCUSSION
In unselected population, the increase in susceptibility was to the extent of 1.91 (LD50 declined from 3.638 to 1.90 μg) and 3.49 fold (LD50 declined from 3.537 to 1.012 μg) to fenvalerate and cypermethrin respectively by the end of fourteenth generation. Selection by pyrethroids increased the level of resistance by 5.24 fold to fenvalerate and 4.91 fold to cypermethrin when the generation advanced to F14.
The second generation of H. armigera did not exhibit any significant variation in developmental and reproductive traits (Tables 1, 2 and 3). The net reproductive rate was 237.437 for unselected population and it was 228.24 and 231.359 for cypermethrin and fenvalerate selected populations respectively. The intrinsic rate of increase was 0.1269 – 0.1274 for pyrethroid selected and 0.1284 for the unselected populations. The duration of the life cycle and doubling time were comparable for both selected and unselected populations.
| Table 1: | Age specific fecundity table for unselected H.armigera (F2) |
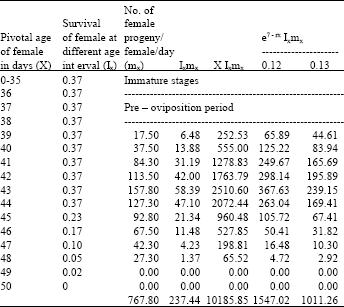 | |
| Intrinsic rate of increase (rm):0.1284, Net reproductive rate (NRR):237.4370, Mean generation time (Tc):42.8992, The capacity for increase (rc) = 0.1275, Finite rate of increase (λ):1.1370, Weekly multiplication rate (WMR): 2.4568, Corrected generation time (T):42.5981, Doubling time (DT): 5.3980 | |
| Table 2: | Age specific fecundity table for fenvalerate selected H.armigera (F2) |
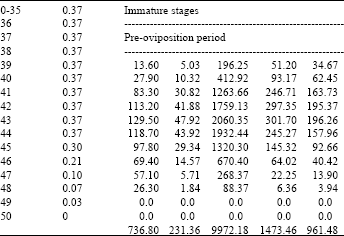 | |
| Intrinsic rate of increase (rm) : 0.1274, Net reproductive rate (NRR): 231.359, Mean generation time (Tc) : 43.1026, The capacity for increase (rc): 0.1263, Finite rate of increase (λ) : 1.1358, Weekly multiplication rate (WMR) : 2.4388 Corrected generation time (T) : 42.7444, Doubling time (DT) : 5.4424 | |
In F7 significant reduction in the net reproductive rate was observed in pyrethroid selected populations (170.867 for fenvalerate selected population; 161.339 for cypermethrin selected population) when compared to the unselected population (213.276) (Tables 4, 5 and 6). Nearly two to three days increase in developmental period over the unselected population was noticed in fenvalerate and cypermethrin selected populations.
| Table 3: | Age specific fecundity table for cypermethrin selected H.armigera (F2) |
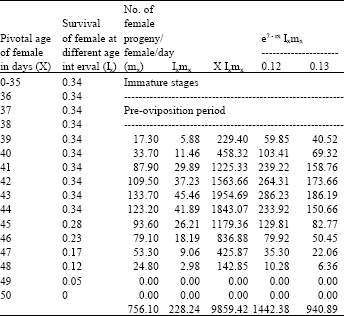 | |
| Intrinsic rate of increase (rm):0.1269, Net reproductive rate (NRR):228.2400, Mean generation time (Tc) : 43.1976, The capacity for increase (rc) : 0.1257, Finite rate of increase (λ) : 1.1353, Weekly multiplication rate (WMR) : 2.4309, Corrected generation time (T) : 42.7944, Doubling time (DT) : 5.4624 | |
| Table 4: | Age specific fecundity table for unselected H.armigera (F7) |
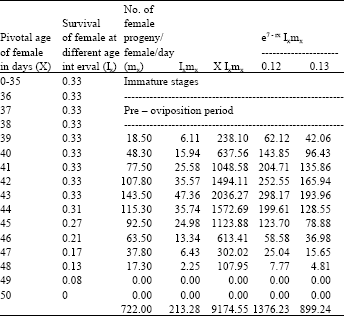 | |
| Intrinsic rate of increase (rm):0.1259, Net reproductive rate (NRR):213.2760, Mean generation time (Tc):43.0173, The capacity for increase (rc):0.1247, Finite rate of increase (λ):1.1341, Weekly multiplication rate (WMR): 2.4134, Corrected generation time (T):42.6067, Doubling time (DT): 5.5072 | |
| Table 5: | Age specific fecundity table for fenvalerate selected H.armigera (F7) |
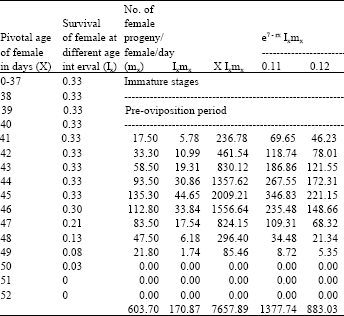 | |
| Intrinsic rate of increase (rm):0.1157, Net reproductive rate (NRR):170.8670, Mean generation time (Tc):44.8179, The capacity for increase (rc):0.1147, Finite rate of increase (λ):1.1226, Weekly multiplication rate (WMR): 2.2474, Corrected generation time (T):44.4394, Doubling time (DT): 5.9918 | |
| Table 6: | Age specific fecundity table for cypermethrin selected H.armigera (F7) |
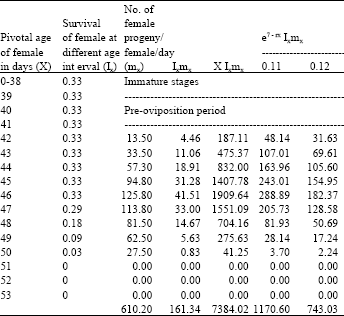 | |
| Intrinsic rate of increase (rm):0.1117, Net reproductive rate (NRR): 161.3390, Mean generation time (Tc):45.7671, The capacity for increase (rc):0.1111, Finite rate of increase (λ):1.1182, Weekly multiplication rate (WMR): 2.1861, Corrected generation time (T):45.4978, Doubling time (DT): 6.2037 | |
| Table 7: | Age specific fecundity table for unselected H.armigera (F14) |
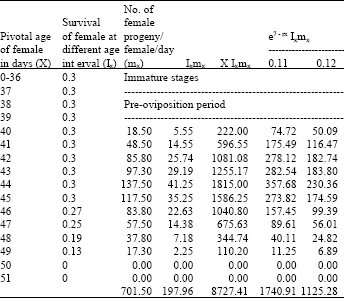 | |
| Intrinsic rate of increase (rm):0.1205, Net reproductive rate (NRR):197.9620, Mean generation time (Tc):44.0863, The capacity for increase (rc):0.1199, Finite rate of increase (λ):1.1280, Weekly multiplication rate (WMR):2.3239, Corrected generation time (T):43.8969, Doubling time (DT): 5.7539 | |
| Table 8: | Age specific fecundity table for fenvalerate selected H.armigera (F14) |
 | |
| Intrinsic rate of increase (rm):0.1079, Net reproductive rate (NRR):123.6770, Mean generation time (Tc):44.9212, The capacity for increase (rc):0.1072, Finite rate of increase (λ):1.1140, Weekly multiplication rate (WMR): 2.1285, Corrected generation time (T):44.6412, Doubling time (DT): 6.4228 | |
Reduction in intrinsic rate of increase, finite rate of increase and weekly multiplication rate of pyrethroid selected populations over the susceptible strain was negligible as they advanced to seventh generation.
| Table 9: | Age specific fecundity table for cypermethrin selected H.armigera (F14) |
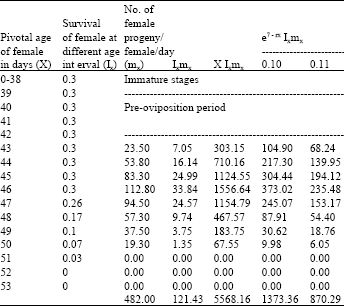 | |
| Intrinsic rate of increase (rm):0.1055, Net reproductive rate (NRR):121.4320, Mean generation time (Tc):45.8541, The capacity for increase (rc):0.1047, Finite rate of increase (λ):1.1113, Weekly multiplication rate (WMR):2.0928, Corrected generation time (T):45.4909, Doubling time (DT): 6.5700 | |
The increase in selection pressure due to pyrethroids led to severe biological deficits. It was manifested through drastic reduction in net reproductive rate in F14. The net reproductive rate dropped down to 123.677 for fenvalerate and 121.432 for cypermethrin. The intense selection pressure might induce the homozygote resistant larvae to divert energy from some area (fecundity) to oxidative metabolism of pyrethroids to defend it. The reduction in the net reproductive rate was not only restricted to pyrethroid selected populations, but also extended to unselected population. However, it was comparatively less in unselected population (Tables 7, 8 and 9). The intrinsic rate of increase, finite rate of increase and the weekly multiplication rate were decreased gradually in selected populations as the generation advanced. The generation time extended by 2.7 days and 1.9 days for cypermethrin and fenvalerate selected populations respectively, whereas in the unselected population the increase was 1.3 days only.
Plapp et al. (1990) suggested that resistant females are less fertile, less fecund, attract fewer males and produce less pheromone. They also observed that resistant females of Heliothis virescens (F.) from the field had mated less than susceptibles and those resistant males from the field responded less to pheromone traps. The early season decline in pyrethroid resistance noted in their monitoring studies was due to reproductive disadvantages allowing rapid replacement of resistant insects by their more reproductively susceptible compatriots. This is in favour of the results obtained in the present investigation. McCutchen et al. (1989) suggested that these reproductive deficits will allow resistance to be managed and without them, resistance could only get worse. So, the presence of reproductive deficit in H.armigera is critical for the long - term success of IRM strategy in India.
ACKNOWLEDGEMENT
The financial support from the Common Fund for Commodities (CFC) Europe, International Cotton Advisory Committee (ICAC) U.S.A. and Natural Resources Institute (NRI), U.K. is acknowledged.
REFERENCES
- Armes, N.J., D.R. Jadhav and D.K.R. Souza, 1996. A survey of insecticide resistance in Helicoverpa armigera in the Indian subcontinent. Bull. Entomol. Res., 86: 499-514.
Direct Link - Birch, L.C., 1948. The intrinsic rate of natural increase of an insect population. J. Anim. Ecol., 17: 15-26.
CrossRefDirect Link - Plapp, F.W., J.A. Jackman, C. Campanhola, R.E. Frisbie, J.B. Graves, R.G. Luttrell, W.F. Kitten and M. Wall, 1990. Monitoring and management of pyrethroid resistance in the tobacco budworm (Lepidoptera: Noctuidae) in texas, mississippi, louisiana, arkansas and oklahoma. J. Economic Entomol., 83: 335-341.