
S. Shahid Shaukat
Not Available
Nadia Munir
Not Available
Imran A. Siddiqui
Not Available
Asian Journal of Plant Sciences
Year: 2003 | Volume: 2 | Issue: 14 | Page No.: 1034-1039







ABSTRACT
The effects of Conyza canadensis (L.) Cronquist on seed germination and early seedling growth of six test plants namely tomato, radish, wheat, corn, millet and mungbean was investigated. Aqueous extract of C. canadensis at different concentrations (25, 50, 75 and 100% stock solution) inhibited the germination, root and shoot growth of all the six test species. Germination was reduced by the shoot extract in the order: tomato > radish > millet = corn > mungbean > wheat. Both root and shoot growth of the test species were reduced to varying degree and shoot growth was usually affected to a greater degree than the root growth, particularly in tomato. Decaying shoot of C. canadensis in sandy-loam at 5, 10 and 20 g/ 400 g soil substantially inhibited germination and seedling growth of bulrush millet (Pennisetum americanum) at all the dosages and no germination occurred at the highest dosage (20 g/400 g soil). Bioassay of the ether extract of C. canadensis disclosed two significant zones of inhibition at Rf values of 0.2-0.3 3 and 0.7-0.8. Chromatography for the phenolics revealed the presence of four phenolic compounds: gallic acid, vanillic acid, catechol and syringic acid.
PDF Abstract XML References Citation
How to cite this article
S. Shahid Shaukat, Nadia Munir and Imran A. Siddiqui, 2003. Allelopathic Responses of Conyza canadensis (L.) Cronquist: A Cosmopolitan Weed. Asian Journal of Plant Sciences, 2: 1034-1039.
DOI: 10.3923/ajps.2003.1034.1039
URL: https://scialert.net/abstract/?doi=ajps.2003.1034.1039
DOI: 10.3923/ajps.2003.1034.1039
URL: https://scialert.net/abstract/?doi=ajps.2003.1034.1039
INTRODUCTION
The vegetational composition and population structure of a site often depends considerably on allelopathic interactions between species (Shaukat et al., 1983). The phenomenon is basically distinct from other mechanisms of interference such as competition, which acts via the depletion of resources (Inderjit and Del Moral, 1997). Allelopathic interactions involve the production and release of chemical substances (allelochemicals) by certain plants that inhibit the growth and development of the individuals of the neighbouring species and sometimes their own (autoallelopathy). Among the weed species reported worldwide, a considerable number are known to possess allelopathic potential (Ashraf and Sen, 1978; Shaukat et al., 1985; Ahmed and Wardle, 1994; Burhan and Shaukat, 1999; Rebaz et al., 2001; Shaukat and Siddiqui, 2002; Tajuddin et al., 2002; Shaukat et al., 2003).
The allelochemicals released from the plants often play a vital role in species sequence and dynamics during secondary succession (Al Saadawi and Rice, 1982; Rice, 1995) and in the determination of spatial pattern and interspecific associations between plant species (Shaukat et al., 1983). Putnam and Weston (1986) listed 90 weed species while Narwal (1994) listed 129 weed species having allelopathic potential. The phenomenon of allelopathy is known to play an important part in weed-weed interaction (Rasmussen and Rice, 1971, Newman and Rovira, 1975; Tajuddin et al., 2002) and weed-crop interaction (Colton and Einhellig, 1980).
Weed-crop interaction with the standpoint of allelopathy has often been explored (Casado, 1995; Inderjit et al., 1996; Lydon et al., 1997; Rajbanshi and Inubushi, 1997; Peres et al., 1998; Ito et al., 1998; Al-Humaid and Warrag, 1998; Tajuddin et al., 2002; Shaukat et al., 2003).
The important allelochemicals include akaloids, terpenoids, flavonoids, steroids, tannins and phenolic compounds that usually have inhibitory effects on crops (Whittaker and Feeny, 1971; Mandava, 1985). Phenolic compounds often constitute the principal allelopathic agents in weeds and other allelopathic plants (Inderjit, 1998; Ferreira et al., 1998; Wang et al., 1998; Burhan and Shaukat, 2000; Shaukat et al., 2003). Once entered into the soil, the bioactive concentration of allelochemicals is determined through their sorption, fixation, leaching and chemical and microbial degradation (Blum, 1999 and Inderjit et al., 1999). Soil chemical, physical and biological characteristics to a great extent are responsible for detoxification or further enhancement of the allelopathic activities of plant diffusates (Cheng, 1995; Schmidt and Ley, 1999). Therefore, it is imperative to perform bioassays in the presence of soil to demonstrate allelopathy of ecological relevance (Foy, 1999).
Conyza canadensis (L.) Cronquist (horse weed: Asteraceae), a cosmopolitan agricultural and ruderal weed, grows abundantly in cultivated and abanodoned fields, lawns, waste grounds and vacant lots in different provinces of Pakistan. Since this weed is often seen to form dense, almost pure stands, it is reasonable to suspect that allelopathy could be involved in the suppression of other plants in the vicinity. Therefore, experiments were designed to test the allelopathic potential of Conyza canadensis. The objectives of this investigation were: 1) to evaluate the effect of aqueous extract of C. canadensis on four crop species in vitro, 2) to study the phytotoxicity of decaying C. canadensis in soil, 3) to examine the phytotoxicity of C. canadensis to tomato in three different soil types and 4) to identify the phenolic principles of C. canadensi.
MATERIALS AND METHODS
Effect of aqueous extract of Conyza canadensis on germination and seedling growth of three test species:C. canadensis was collected from the experimental field of Crop Disease Research Institute, Karachi University Campus and its shoot extract was examined for possible allelopathic activity. The plant material was air-dried under shade and chopped into small pieces. Extract of C. canadensis was prepared by soaking 10 g plant material in 100 ml of distilled water for 24 h to obtain stock solution. Using stock solution (100%), three other concentrations were prepared i.e, 25, 50 and 75%. Effect of various concentrations was tested against tomato (Lycopersicon esculentum Mill.), radish (Raphanus sativus L.), bulrush millet (Pennisetum americanum (L.) Schumann), corn (Zea mays L.), mungbean (Vigna radiata (L.) Wilczek) and wheat (Triticum aestivum L.).
Crop seeds were first surface sterilized by 0.3% calcium hypochlorite for 5 min and then placed on 9 cm diam., sterile Petri plats on two layers of Whatman No. I filter paper. Each plate received 5 ml of the extract. Distilled water was used for controls. Final germination was noted at 96 h and shoot and root length of the seedlings were measured.
Effect of decaying Conyza canadensis on germination and seedling growth of Pennisetum americanum: Dried chopped material of C. canadensis was mixed thoroughly with loamy sand (80% sand, 12% silt and 8% clay) at 10, 15 and 20 g/400 g soil. Pots were watered once and soil was left for biodegradation. After one week, 10 seeds of bulrush millet (Pennisetum americanum(L.) Schumann) were sown in each pot. Controls and treatments were replicated four times and pots were randomized in blocks on a greenhouse bench. Final emergence percentage was recorded while shoot and root lengths were measured after one week.
Effect of Conyza canadensis on germination and seedling growth of Pennisetum americanum in three soil types: Dried powdered shoot of Conyza canadensis was thoroughly mixed at a rate of 20 g/400 g of three different soil types including a sandy soil (86% sand, 8% silt and 6% clay), a loamy sand (80% sand, 12% silt and 8% clay) and a sandy loam (68%, 17% silt, 15% clay). A small quantity of farmyard manure was sprinkled in all pots. Ten seeds of bulrush millet (Pennisetum americanum (L.) Schumann) were sown in each pot. The pots were randomized on a laboratory bench in blocks and were watered daily. Controls and treatments were replicated four times and pots were randomized within blocks on a greenhouse bench. Final emergence percentage and shoot and root lengths were measured after one week.
Bioassay: Ten-gram air-dried shoot of C. canadensis were blended in 200 ml distilled water. The centrifuged homogenate was adjusted to pH 3 with 0.5 N H2SO4 extracted thrice with peroxidase-free ether and evaporated to dryness using argon gas. Two ml of 80% ethanol was added to the dried material and was streaked on Whatman No. I filter paper. Duplicate 10 cm wide chromatograms were developed by descending chromatography in isopropanol: ammonia: water (10:1:1,v/v/v). When the solvent had moved 30 cm, the chromatograms were dried and 10 equal width strips were cut and assayed for growth regulators using wheat coleoptile straight growth test of Nitsch and Nitsch (1956). Five mm segments of 3 day old dark grown wheat were excised and floated in distilled water for I h. Ten coleoptile segments were placed in between two strips of the same Rƒ value and kept in 11.5 cm diam. Petri plates over two layers of tissue paper moistened with 4 ml 0.02 M citrate phosphate buffer (pH 4.8). After 48 h of growth in dark, the length of coleoptile segments was measured.
Chromatography: Ether extract of C. canadensis was evaporated to dryness, dissolved in 2 ml of 80% ethanol and used for loading on silica gel F254 thin layer chromatographic plate. The chromatogram was developed in acetic acid-chloroform 1:9 v/v by ascending chromatography using reference phenolic 'compounds. Phenolic compounds were detected using ferric chloride-ferric cyanide reagent and UV light (Harborne, 1973).
Statistical analysis: Data sets were subjected to analysis of variance (ANOVA) of factorial analysis of variance (FANOVA) depending upon experimental design. Percentage data were arcsine transformed prior to analysis (Sokal and Rohlf, 1995). As a follow up of ANOVA or FANOVA, Fisher's a least significant difference (LSD) test and Duncan's multiple range test were performed at p=0.05. Percentage germination or emergence data were arcsine transformed in order to conform to normality assumption. In data sets where significant heteroscedasticity occurred, log(x)-transformed values were used in the analysis of variance. Computer programs for the analyses were developed in Microsoft FORTRAN-77 by the first author (S. S. S.).
RESULTS
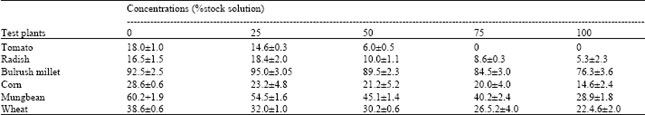
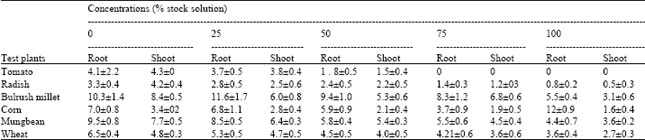
Effect of aqueous extract on germination and seedling growth of the test species: Germination of all the six test species was inhibited by different concentrations of the extract (p < 0.01) over the controls (Table 1). The inhibitory effect on germination increased with the increase in concentration. Different species were affected to a different extent; the degree of inhibition varied in the order: tomato > radish > corn = mungbean > wheat > bulrush millet. Similarly, seedling growth of the test species was also adversely affected and growth reduction was greater at higher concentrations. Root and shoot growth were more or less equally affected by C. canadensis extract; they were inhibited in the order: tomato > radish > corn = wheat > mungbean > millet (Table 2).
Phytotoxicity of decaying C. canadensis: Emergence percentage of bulrush millet was significantly (p < 0.001) suppressed at 15 and 20 g/400 g soil the dosages of decaying C. canadensis (Table 3). Emergence percentage declined sharply with the doubling of the dosage. Likewise, both root and shoot growth were significantly (p<0.001) retarded at both the higher dosages of the decaying shoot material compared to the controls.
| Table 1: | Effect of aqueous shoot extract of Conyza canadensis on final germination percentage of six test plants |
 | |
| Table 2: | Effect of aqueous shoot extract of Conyza canadensis on shoot and root growth (cm) of six test plants |
 | |
| Table 3: | Phytotoxic effects of decaying Conyza canadensis on emergence and root and shoot growth of bulrush millet (Pennisetum americanum) |
 | |
| Table 4: | Phytotoxic effects of decaying Conyza canadensis on emergence and root and shoot growth of bulrush millet (Pennisetum americanum) |
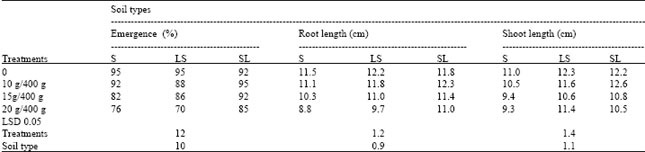 | |
| S = sandy soil; LS = loamy sand; SL = sandy loam | |
| Table 5: | Rƒ-values (x 100) of phenolic principles in ether fraction of aqueous extract of Conyza canadensis and their reaction to a developing reagent and UV light |
 | |
This inhibitory effect was more pronounced at the dosage of 20 g shoot material/400 g soil.
Effect of Conyza canadensis on germination and seedling growth of Pennisetum americanum in three soil types: Soil type had a significant influence on the activity of Conyza canadensis shoot powder (p < 0.01) (Table 4). The emergence percentage was most affected by C. canadensis amendment in sandy soil followed by loamy sand and sandy loam. Likewise, root and shoot growth was suppressed by C. canadensis to a greater degree in sandy soil compared to sandy loam and loamy sand. Root and shoot growths were more or less equally affected by the amendments. In all cases suppressive effect occurred only at 15 and 20 g/400 soil and no significant toxic effect was observed at l0g/400g soil.
Wheat coleoptile bioassy: Wheat coleoptile bioassay disclosed two significant inhibitors at Rƒ-values of 0.2-0.3 and 0.7-0.8, while one significant promoter was detected at Rƒ-values of 0.4-0.5.
Chromatographic study: Chromatograms sprayed with ferric chloride-ferricyanide reagent or examined under UV light, disclosed four spots, which were matched with the standard phenolic, compounds and identified as gallic acid, vanillic acid, catechol and syringic acid (Table 5).
DISCUSSION
This study provides evidence on the allelopathic potential of C. canadensis. Seed germination and seedling growth of tomato, radish, wheat, corn, bulrush millet, mungbean and wheat were inhibited to varying degree by aqueous extract of C. canadensis. Extracts of a number of weed species have been shown to be inhibitory to germination of crop seeds (Shaukat et al., 1985; Rafique and Hafeez, 1994; Casado, 1995; Hoffmann et al., 1996; Demchuk and Yurchak 1996; Lydon et al., 1997; Inderjit et al., 1998). To the best of our knowledge there are no previous report on the allelopathic potential of C. candensis. The differential effect of the aqueous extract of C. canadensis on germination and early seedling growth of the six test species presumably because the different phenolic compounds (and possibly other inhibitors) might have variable effects due to morphological and/or physiological differences among the test species.
Decaying C. canadensis in loamy sand caused marked inhibition of germination and seedling growth of bulrush millet. Wilson and Rice (1968) have reported both stimulatory and inhibitory effect on various crop species with decaying sunflower leaves. In the present study, maximum reduction was observed in soil incorporated with 20 g shoot material. This may presumably be due to the release of phytotoxins from the decaying C. canadensis that remain active and stable for considerable duration in soil. Due to the presence of lesser quantity of colloidal material in the soil, the adsorption of phytotoxins released during the process of decay is expected to be minimal. Likewise, Shaukat et al. (1985) reported adverse effects of decaying Citrullus colocynthes while Burhan and Shaukat (1999) reported inhibitory effects of decaying Argemone mexicana on bulrush millet. Shaukat et al. (2003) also observed deleterious effects of decaying Launaea procumbens on germination and growth of millet.
The phytotoxicity of C. canadensis varied in the three soil types that differed in soil texture. For example, the greatest activity was recorded in sandy soil. This is presumably due to the fact that this soil has a very small proportion of colloidal material (least clay content) and hence the adsorption of the allelochemicals is minimal. Also, because the microbial or chemical degradation of the phytotoxic compounds is supposed to be least in such soil. Although leaching of chemicals is expected to be relatively a greater rate in sandy soil but still this would take some time, on the other hand, the phytotoxicity to the seedlings was noted only one week after the soil amendment.
It is anticipated that the phytotoxins originating from C. canadensis through leaching, decay or exudation from roots, would eventually accumulate in the soil in biologically significant amounts and consequently playing an active role as a habitat factor, exerting a causative influence on growth and development of other plants in the neighbourhood. The wheat coleoptile bioassay of C. canadensis disclosed two inhibitory zones that presumably represented compounds of phenolic nature. In addition to the two inhibitors, a promoter was also detected. The latter was presumably a phytohormone. Thin layer chromatography revealed the presence of four phenolic compounds including vanillic acid, catechol, gallic acid and syringic acid. Regarding the chemical nature of the phytotoxins, attention was focused on the most common allelochernicals the phenolic compounds. However, the occurrence of phytotoxic chemicals other than phenolic compounds can not be ruled out that may constitute important allelopathic agents. The toxic nature of phenolic compounds with reference to plants have been reported by several workers (Stowe et al., 1987; Blum, 1996; Inderjit, 1998; Burhan and Shaukat, 2000).
The results presented in this paper lead to the conclusion that C. canadensis is considerably phytotoxic and often being a dominant species in some of the weed communities in Southern Sindh accumulates phytotoxins over a period of several years that can eventually suppress the associated crops and alter the weed community structure. Thus, the populations of this weed should be carefully monitored and managed, particularly in vulnerable crops such as tomato and radish.
ACKNOWLEDGMENTS
This research was supported by a grant from Dean Faculty of Science, University of Karachi, which is gratefully acknowledged. Technical assistance of Maria Hamid, University of Karachi and S. Azhar Ali, Haider Farms, Gharo is greatly appreciated.
REFERENCES
- Ahmed, M. and D.A. Wardle, 1994. Allelopathic potential of vegetative and flowering ragwort (Senscio jacobaea L.) plants against associated Pasture species. Plant Soil, 164: 61-68.
CrossRef - Humaida, I.A. and A. Warrag, 1998. Allelopathic effects of mesquite (Prosopis juliflora) foliage on seed germination and seedling growth of bermudagrass (Cynodon dactylon). J. Arid Environ., 38: 237-244.
CrossRef - Burhan, N. and S.S. Shaukat, 1999. Allelopathic potential of Argemone mexicana L. A tropical weed. Pak. J. Biol. Sci., 2: 1268-1273.
CrossRefDirect Link - Inderjit, 1998. Influence of Pluchea lanceolata (Asteraceae) on selected soil properties. Am. J. Bot., 85: 64-69.
Direct Link - Ito, I., K. Kobayashi and T.Y. Yoneyama, 1998. Fate of dehydrometricaria ester added to soil and its implications for the allelopathic effect of Solidago altissima L. Ann. Bot., 82: 625-630.
CrossRef - Rasmussen, J.A. and E.L. Rice, 1971. Allelopathic effects of Sporobolus pyramidatus on vegetational patterning. Am. Midland Naturalist, 86: 309-326.
Direct Link - Rebaz, Z., S.S. Shaukat and I.A. Siddiqui, 2001. Allelopathic potential of Anagallis arvensis L.: A cosmopolitan weed. Pak. J. Biol. Sci., 4: 446-450.
CrossRefDirect Link - Shaukat, S.S., D. Khan and S.T. Ali, 1983. Suppression of herbs by Inula grantioides Bioss. in the Sindh Desert, Pakistan. Pak. J. Bot., 15: 43-67.
Direct Link - Shaukat, S.S., T. Zamarrud and A. Imran-Siddiqui, 2003. Allelopathic potential of Launaea procumbens (Roxb.) rammaya and rajgopal: A tropical weed. Pak. J. Biol. Sci., 6: 225-230.
CrossRefDirect Link - Stowe, L.G., B.S. Kil and Y.J. Yim, 1987. A survey of old-field herbs for susceptibility to phenolic compounds. Korean J. Bot., 30: 11-20.
Direct Link - Tajuddin, Z., S.S. Shaukat and A.I. Siddiqui, 2002. Allelopathic potential of Solanum forskalii Dunal: A tropical ruderal weed. Pak. J. Biol. Sci., 5: 866-868.
CrossRefDirect Link - Wang, M., J. Li, M. Rangarajan, Y. Shao, E.J. LaVoie, T.C. Huang and C.T. Ho, 1998. Antioxidative phenolic compounds from sage (Salvia officinalis). J. Agric. Food Chem., 46: 4869-4873.
CrossRefDirect Link - Whittaker, R.H. and P.P. Feeny, 1971. Allelochemics: Chemical interactions between species. Science, 171: 757-770.
CrossRefDirect Link - Wilson, R.E. and E.L. Rice, 1968. Allelopathy as expressed by Helianthus annus L. and its role in old-field succession. Bull. Torrey Bot. Club, 95: 432-448.
Direct Link - Ashraf, N. and D.N. Sen, 1978. Allelopathic potential of Celosia argentea in and land crop field. Oecol. Plant., 13: 331-338.
Direct Link - Burhan, N. and S.S. Shaukat, 2000. Effect of phenolic compounds on germination and seedling growth of some crop plants. Pak. J. Biol. Sci., 3: 269-274.
Direct Link - Inderjit, C.L. Foy and K.M.M. Dakshini, 1998. Pluchea lanceolata: A noxious perennial weed. Weed Technol., 12: 190-193.
Direct Link - Blum, U., 1996. Allelopathic interactions involving phenolic acids. J. Nematol., 28: 259-267.
Direct Link