
Mohammad Subhan
Not Available
M. Qasim
Not Available
Khalil Ahmad
Not Available
Asian Journal of Plant Sciences
Year: 2002 | Volume: 1 | Issue: 3 | Page No.: 257-260







ABSTRACT
An 8 x 8 diallel analysis study on cotton (Gossypium hirsutum L.) was conducted at Faculty of Agriculture, Gomal University, D. I. Khan, during, 1996-99 to determine the type of gene action in the control and expression of some important quantitative characters like seed index and lint index in F1 and as well as in F2 generations. The analysis of variance exposed that differences among genotypes for the characters mentioned were highly significant. The Hayman-Jinks model proved to be partial adequate for the characters mentioned in F1 while completely adequate in the F2 generations in this manuscript.
PDF Abstract XML References Citation
How to cite this article
Mohammad Subhan, M. Qasim and Khalil Ahmad, 2002. Estimates
of Gene Effects for Some Quantitative Characters in Upland cotton (Gossypium
hirsutum L.). Asian Journal of Plant Sciences, 1: 257-260.
DOI: 10.3923/ajps.2002.257.260
URL: https://scialert.net/abstract/?doi=ajps.2002.257.260
DOI: 10.3923/ajps.2002.257.260
URL: https://scialert.net/abstract/?doi=ajps.2002.257.260
INTRODUCTION
The cotton (Gossypium hirsutun L.) being biggest foreign exchange earner in Pakistan, attracts utmost attention of plant breeders to evolve high yield varieties. Consequently, Pakistan has become one of the top cotton producing countries in the world. Abundance of literature is available on practical usefulness of diallel analysis to understand type of gene action and genetic potential of parents in cotton like Singh et al. (1985), Raza et al. (1990), Murtaza et al. (1992), Azhar et al. (1994), Busharat et al. (1998 a). However results of these studies vary as concluded by Rehman et al. (1988), Busharat et al. (1998 b) and Subhan et al. (2000), depending on genotypes and environmental conditions. This study is aimed to employ diallel cross analysis in understanding breeding utility of eight cotton cultivars for different economic traits.
MATERIALS AND METHODS
The F1 and F2 populations were developed at Faculty of Agriculture, Gomal University, D.I.Khan, during, 1996-99 by crossing eight cotton cultivars viz. CYTO 9/91, B-496, SLS-1, Niab-78, NIAB-313/12, B-622 and Niab-78, NIAB-313/12, B-622, NIAB-92 and CYTO-11/91 in a diallel fashion during the months of August and September, 1996. The data of F1 and F2 obtained from the crosses and their selfed parents grown in Randomized Complete Block Design with three replications. Each plot comprised single 3-m long row with 30 cm plant to plant and 75 cm row to row distance. The recommended production package was adopted. The data on individual plant basis were recorded at crop maturity and statistically analyzed for seed and lint indices through diallel technique developed by Hayman, (1954a) and applied by Mather and Jinks (1977).
RESULTS AND DISCUSSION
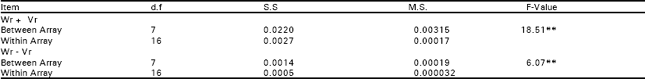
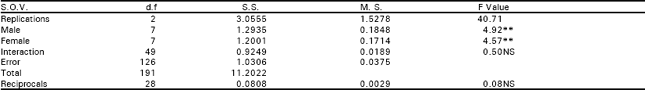
The analysis of variance depicted that differences among the genotypes were highly significant for seed index and lint index both in F1 and F2 generations. This reflected presence of adequate genetic variability which could be Table exploited in different crossing programs (Table 1). Additive genetic variation seemed to exist for these traits in both the generations as the mean squares due to both male and female parents were highly significant (P<0.01). The interaction differences among male and female parents for seed index and lint index were also significant indicating the presence of non additive heritable genes while the reciprocal differences were recorded as non significant exposing nothing like maternal effects in F1 generation.
| Table 1: | Estimates of mean squares and F ratios of different quantitative traits of cotton (Gossypium hirsutum L.) in F1 and F2 generation. |
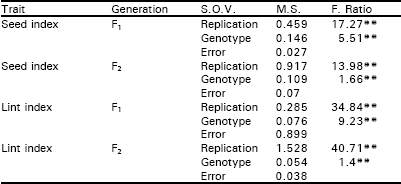 | |
| **, Highly significant | |
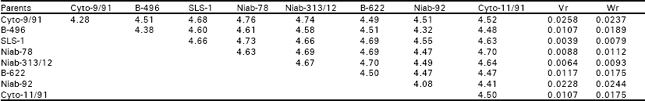
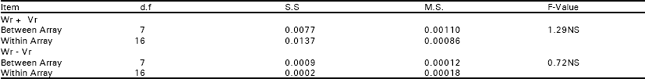
But the interaction and reciprocal differences for the same mentioned characters were noted as non significant depicting absence of non additive heritable genes and maternal effects in F2 generations (Table 2a, 3a, 4a, 5a). To test the adequacy of additive dominance model of the data, regression analysis was carried out. The analysis of the data (Table 2b and 3b) revealed that the value of regression coefficient b (b=0.911±0.158, b=0.688±0.129, b=1.021±0.071, b=0.729±0.1326) differed significantly from zero and were none significantly deviated from unity for both the traits i.e., seed index and lint index in both the generations (Table 2b, 3b, 4b, 5b). The results of analysis of variance for arrays showed that Wr-Vr did differ significantly between the arrays in seed and lint indices in their F values which are enough to mold the model partially adequate regarding the characters mentioned in F1 population. This also indicated the participation of epistacy or multiple allelism in F1 generation (Table 2c, 3c). The non-significant differences for Wr+Vr indicated the absence of dominance genetic effects, whereas the non-significant differences for Wr-Vr reflected the absence of non-allelic interaction in case of seed and lint indices. The above two tests disclosed that the additive dominance model was fully adequate for the characters mentioned in F2 population (Table 4c, 5c). The graphic representation of Wr/Vr (Fig. 1 and 2) highlighted that Cyto-11/91 and B-622 parents were exposed with maximum dominant genes, while Niab-92 with maximum recessive genes for seed indices (F1, F2) respectively.
| Table 2a: | Analysis of variance of F1 of diallel data for seed index |
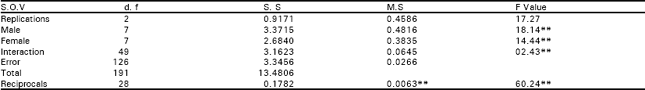 | |
| NS, non-significant; *, significant (P<0.05) and **, highly significant (P<0.01) | |
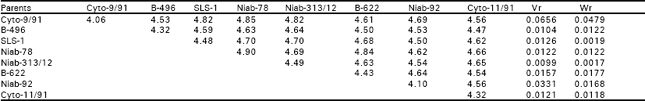
| Table 2b: | Mean data over replications and reciprocals |
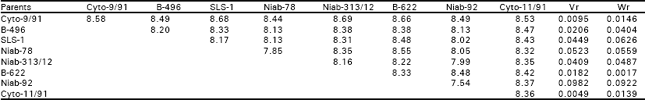 | |
| Regression coefficient (b) = 0.9110±0.1580, difference of b from zero (b0) = 5.7659** different for b from unity (b1) =0.5631NS NS, non-significant; **, highly significant | |
| Table 2c: | Analysis of variance for arrays |
 | |
| **, highly significant; *, significant | |
| Table 3a: | Analysis of variance of F1 of diallel data for lint index. |
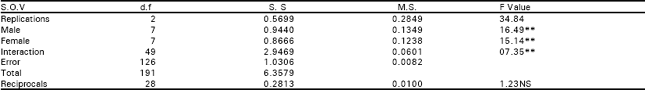 | |
| NS, non-significant, *, significant (P<0.05) and **, highly significant (P<0.01) | |
| Table 3b: | Mean data over replications and reciprocals. |
 | |
| Regression coefficient (b) = 0.6878±0.1285, difference of b from zero (b0) = 5.3519** different for b from unity (b1) =2.4291 NS, NS, non-significant; **, highly significant | |
| Table 3c: | Analysis of variance for arrays |
 | |
| **, highly significant | |
| Table 4a: | Analysis of variance of F2 of diallel data for seed index. |
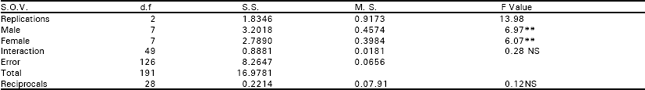 | |
| NS, non-significant; *, significant (P<0.05); **, highly significant (P<0.01) | |
| Table 4b: | :Mean data over replications and reciprocals |
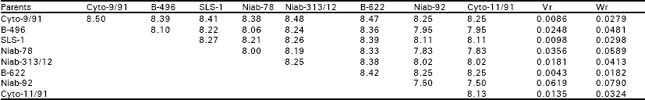 | |
| Table 4c: | Analysis of variance for arrays |
 | |
| **, highly significant; NS, non-significant | |
| Table 5a: | Analysis of variance of F2 of diallel data for lint index |
 | |
| NS, non-significant, *, significant (P<0.05) and **, highly significant (P<0.01) | |
| Table 5b: | Mean data over replications and reciprocals. |
 | |
| Table 5c: | Analysis of variance for arrays |
 | |
| NS, non-significant | |
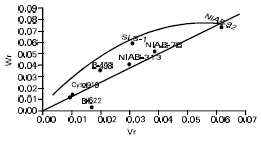 | |
| Fig. 1: | Wr/Vr graph for seed index (F1) |
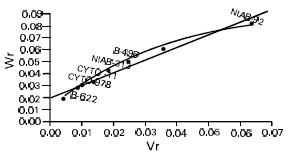 | |
| Fig. 2: | Wr/Vr graph for seed index (F2) |
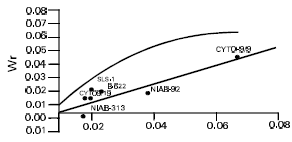 | |
| Fig. 3: | Wr/Vr graph for lint index (F1) |
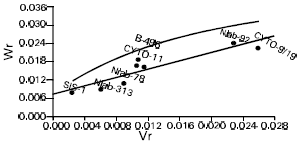 | |
| Fig. 4: | Wr/Vr graph for lint index (F2) |
In case of lint indices (F1, F2) Niab-313/12 and SLS-1 were recorded with maximum dominant genes, though Cyto-9/91 was noted with maximum recessive genes (Fig. 3 and 4). The regression line cut the Wr axis through the point of intersection thus reflected complete dominance for seed and lint indices in F1 population (Fig. 1 and 3). Furthermore, the regression line cut Wr axis above the origin, therefore, signified the presence of additive dominance gene action for the characters like seed and lint indices in F2 population (Fig. 2 and 4). The estimate of components of genetic variation showed that both additive and non-additive genetic effects appeared to be important in the inheritance mechanism of these traits. However after the comparison of these two components, it was depicted that genes acting additively, were more pronounced than genes acting non-additively. Our results are in confirmative with those of earlier workers, Raza et al. (1990), Rehman et al. (1993), Azhar et al. (1994), Busharat et al. (1998a), Busharat et al. (1998b), Subhan et al. (2000). However finding of some authors, Rehman et al. (1988), Azhar and Rana (1993) and Khan et al. (1994) don’t agree to these observations. Their differences might largely be attributed to different experimental material with a varied genetic back ground in this respect. The Hayman-Jinks model proved to be partial adequate for seed and lint indices in F1 generation while both the characters are completely adequate in F2 generation in this exploration. This type of gene action i.e., additive type with partial dominance, indicates signs for isolation of new hybrids from the lint point of view.
REFERENCES
- Azhar, F.M., N. Khan and S.K. Ajmal, 1994. Genetic basis of variation in upland cotton. J. Agric. Res., 32: 9-16.
Direct Link - Singh, M., T.H. Singh and G.S. Chahal, 1985. Genetic analysis of some seed quality characters in upland cotton (Gossypium hirsutum L.). Theoret. Applied Genet., 71: 126-128.
CrossRefDirect Link - Subhan, M., H.U. Khan and R.D. Ahmad, 2000. Comparison of the gene action controlling metric characters in upland cotton (Gossypium hirsutum L.). Pak. J. Biol. Sci., 3: 2087-2090.
CrossRefDirect Link