
Assion Sétu Mivedor
Laboratory of Plant Virology and Biotechnology, Higher School of Agronomy, University of Lomé, Lomé, Togo
Kodjovi Atassé Dansou-Kodjo
Laboratory of Plant Virology and Biotechnology, Higher School of Agronomy, University of Lomé, Lomé, Togo
Djodji Kossikouma Adjata
Laboratory of Plant Virology and Biotechnology, Higher School of Agronomy, University of Lomé, Lomé, Togo
LiveDNA: 228.5838
Justin Simon Pita
Laboratory of Plant Virology, Université Félix Houphouët-Boigny, Pôle Scientifique et d'Innovation, Bingerville, Côte d'Ivoire, BP V34, Abidjan
Asian Journal of Plant Pathology
Year: 2020 | Volume: 14 | Issue: 1 | Page No.: 11-20







ABSTRACT
Background and Objective: Cassava (Manihot esculenta, Crantz) is one of the most important staple food crops in Togo after maize. Cassava mosaic disease (CMD) is transmitted either through infected cuttings or by whiteflies. This study was to evaluate the incidence of begomoviruses causing CMD in the country’s cassava fields and updating knowledge of these viruses could help to better manage CMD in Togo. Materials and Methods: An initial investigation of viruses which might account for these symptoms was carried out using symptomatic as well as asymptomatic samples collected during epidemiological surveys performed in 2015 (472 samples) and 2017 (300 samples). Virus identification was carried out using PCR. Results: The presence of two strains of cassava mosaic begomoviruses (CMBs): African cassava mosaic virus (ACMV) and East African cassava mosaic virus (EACMV). The CMB incidence was lower in 2017 (12.42%) than in 2015 (26.23%) and ACMV was the dominant CMB across the regions sampled. Conclusion: The results indicate that good cultural practices and introduction of resistant cultivars, together with continuous disease monitoring, are required to reduce the impact of CMD in Togo.
PDF Abstract XML References Citation
How to cite this article
Assion Sétu Mivedor, Kodjovi Atassé Dansou-Kodjo, Djodji Kossikouma Adjata and Justin Simon Pita, 2020. Identification and Incidence of Cassava Mosaic Begomoviruses in Togo. Asian Journal of Plant Pathology, 14: 11-20.
URL: https://scialert.net/abstract/?doi=ajppaj.2020.11.20
URL: https://scialert.net/abstract/?doi=ajppaj.2020.11.20
INTRODUCTION
Cassava (Manihot esculenta Crantz) is a staple crop in sub-Saharan Africa, where it provides food security. In Togo, it is one of the most consumed foodstuffs, with an annual yield1 of 1.041.682 Mt. Regions of high cassava production are Maritime (37%) and Plateaus (33%), followed by Central and Kara with shares of national production of 25 and 5%, respectively (Fig. 1).
In recent years, Savannah, traditionally a non-cassava growing region, has cultivated this crop in several areas, this interest was triggered by the nutritional and economic prospects of the manufactured cassava product ‘garri’3. Cassava production in Africa is severely constrained by pests and diseases that cause high yield losses4. Among these, cassava mosaic disease (CMD), which occurs wherever cassava is grown, is the most economically-damaging disease of cassava5. The disease is caused by cassava mosaic begomoviruses (CMBs) (Family Geminiviridae; Genus Begomovirus) transmitted by the whitefly vector Bemisia tabaci (Gennadius) and through cassava cuttings used routinely for vegetative propagation6. The viruses that cause CMD in sub-Saharan Africa include East African cassava mosaic Cameroon virus, East African cassava mosaic Malawi virus, East African cassava mosaic Zanzibar virus, Southern Africa cassava mosaic virus and those found in Togo, namely East African cassava mosaic virus (EACMV), African cassava mosaic virus (ACMV) and Indian cassava mosaic virus (ICMV)7. Crop losses by ACMV alone can amount to 30-40% and EACMV can cause losses of up to 68%. In case of ACMV-EACMV co-infection, losses can reach 82% in Africa8. In recent years, CMD caused by EACMV in East Africa has rapidly spread to several West African countries such as Burkina Faso and Togo9.
In Togo, viruses responsible for CMD were identified in previous studies as ACMV, EACMV and ICMV. Cultivation of cassava in Togo presents a challenge because all cultivars grown in the country are susceptible to CMD10. This susceptibility to CMD leads to low productivity for which there may be several underlying causes: large-scale non-adoption of good agricultural practices, unimproved planting and the lack of phytosanitary knowledge concerning control of pests and diseases11. Other factors contributing to difficulties in disease management include farmers using CMD-infected cuttings as planting material and possible variation in the degree of CMD susceptibility among undocumented cassava varieties grown in a given field.
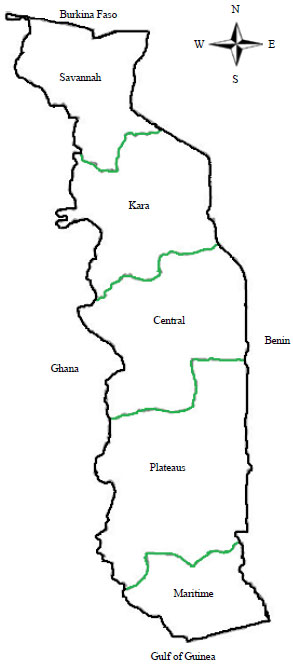 | |
| Fig. 1: | Agro-ecological regions of Togo |
| Source: Author’s work (Modified from Leclerc2) | |
However, there are local varieties which, despite exhibiting disease symptoms, have been selected for planting by farmers because they have greater tuber yield than other local cultivars12. In addition to these preferred local varieties, farmers also plant improved cultivars with high yield potential such as Gbazé-kouté, TMS 96/409, TMS 92/0326 and TMS 95/01665.
The aim of this preliminary study was to evaluate the incidence of begomoviruses causing CMD in the country’s cassava fields. Updating knowledge of these viruses could help to better manage CMD in Togo.
MATERIALS AND METHODS
Sample collection: A harmonized field sampling protocol was adopted by the West African Virus Epidemiology (WAVE) project, which includes Togo and was based on previously described methods13. Cassava fields surveyed were 10 km apart and, for each farm surveyed, 30 plants were assessed randomly along two diagonals. In both survey years, the geolocation coordinates of fields were recorded using a global positioning device (Garmin Dakota TM 10).
Samples of cassava were collected and associated weeds from farms located across the five cassava-producing agro-ecological regions of Togo in atwo-stage survey process: 2015 and 2017. Weeds (uncultivated plants) were sampled also as they can act as intermediate host plants for virus vectors when cassava (their preferred host) is unavailable14. In 2015, data were collected using hardcopy survey forms at the farm locations (Fig. 2).
In 2017, data collection process was refined, in collaboration with the University of Cambridge and Rothamsted Research in the UK and converted our data collection form into a specifically designed application that was uploaded onto a tablet. Locations sampled in 2017 are shown in Fig. 3.
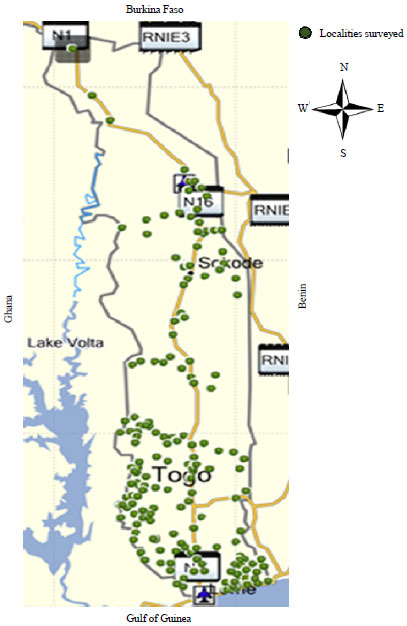 | |
| Fig. 2: | Cassava farms sampled in 2015 |
Source: Author’s work, N1: National road 1, Lome-Cinkansé, RNIE 3: National highway of Benin, It passes from North to South in the West of the country running near the border with Togo, N16: National road 16, Kara-Kétao axis leads to the Benin-Togo border to the East | |
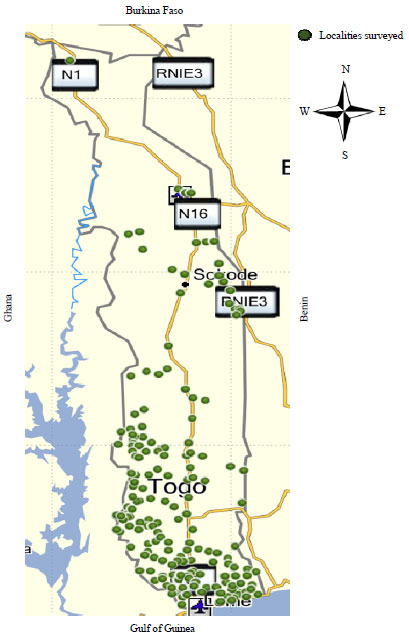 | |
| Fig. 3: | Cassava farms sampled in 2017 |
Source: Author’s work, N1: National Road 1, Lome-Cinkansé, RNIE3: National highway of Benin, it passes from North to South in the West of the country running near the border with Togo, N16: National Road 16, Kara-Kétao axis leads to the Benin-Togo border to the East | |
Cassava leaf and weed samples were labeled, dried, pressed and stored at 25°C (laboratory temperature) prior to analysis. Field assessment of CMD symptom severity on samples of cassava leaf and weeds was carried out using a scale of 1-5. 1: No symptoms (asymptomatic), 2: Mild, 3: Moderate, 4: Severe and 5: Very severe15. The samples collected were representative of all levels of disease severity within a field.
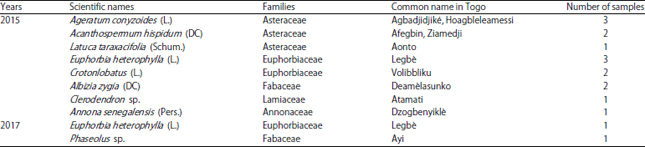
In 2015, we collected 472 samples: 191 asymptomatic and 266 symptomatic samples of cassava leaf and 15 symptomatic samples of 8 species of weed growing in the same cassava fields (Table 1). In 2017, we collected 300 samples: 213 asymptomatic and 85 symptomatic samples of cassava leaf and two of weeds.
DNA extraction and detection of CMD viruses: The DNA was extracted from cassava and weed leaf samples using a modified cetyltrimethylammonium bromide (CTAB) method16. For the 2015 samples, PCR identification of begomoviruses based on published protocols17 was carried out using two pairs of specific primers (JSP001/JSP002 and JSP001/JSP003) to amplify the coat protein (CP) region of component DNA-A (Table 2).
For the 2017 samples also guided by published protocols17,18 and used three pairs of specific primers to target the following DNA-A regions: CP, AL1 and AR2 (Table 3). The ACMV-AL1/F and ACMV-AR0/R primers were used to amplify samples that could not be detected by the other 2 primer sets.
| Table 1: | Weed samples collected in 2015 and 2017 |
 | |
| Table 2: | Primer pairs used for processing 2015 samples |
 | |
| ACMV: African cassava mosaic virus, EACMV: East African cassava mosaic virus, CP: Coat protein | |
| Table 3: | Primer pairs used for processing 2017 samples |
 | |
| ACMV: African cassava mosaic virus, EACMV: East African cassava mosaic virus, CP: Coat protein | |
The PCR parameters were optimized for 25 μL reactions using the following component concentrations: 10 mM dNTP, 1×Taq DNA polymerase buffer, 10 mM MgCl2, 0.8 units Taq DNA polymerase, 10 μM of each complementary and virus-sense primers and 1 μL of DNA template. An Applied Biosystems Thermal Cycler was used (Applied Biosystems, CA, USA). The reaction cycles were 94°C for 5 min followed by 35 cycles at 94°C for 45 sec, 55°C for 45 sec, 72°C for 55 sec and a final extension at 72°C for 7 min. The PCR products were electrophoresed at 100 V for 45 min on a 1% agarose gel stained with ethidium bromide (10 mg mL–1) alongside a 1.0kb DNA ladder. Bands were visualized using a UV transilluminator (TFX-20.MC, Vilbert Lourmat, Marne la Vallée, France). For both 2015 and 2017 samples, virus incidence was calculated as the proportion of samples in which begomoviruses were detected. In this study, the only molecular investigation on the samples was through the use of PCR and virus-specific primers.
RESULTS
Detection of CMBs: The PCR output indicated the presence of two begomoviruses, ACMV (Fig. 4) and EACMV (Fig. 5), based on their expected band size.
Begomovirus incidence in 2015: Of the 472 cassava leaf and weed samples analyzed for CMD, 27.54% tested positive for ACMV (N = 130) and 2.11% for EACMV (n =10). Among the 266 symptomatic samples, ACMV incidence was 48.49%, EACMV was 3.38% and mixed ACMV-EACMV infection was 2.25%. Virus incidence in asymptomatic cassava was low (<0.52%) and undetectable in weeds (Table 4). Severity descriptions used in the Table 4 headings, namely moderate, severe and very severe, refer to severity scores 3, 4 and 5, respectively.
Begomovirus incidence in 2017: Among the 300 samples of cassava and weed material analyzed, 12% tested positive for ACMV (n = 36), 1.33% tested positive for EACMV (n = 4) and 0.33% (n = 1) for mixed ACMV-EACMV infection (Table 5). Among the 85 symptomatic cassava samples, the incidence of sole ACMV and EACMV infection was 30.58 and 4.70%, respectively; and among the 213 samples of asymptomatic cassava, incidence of ACMV as a single infection was 4.69%. Severity descriptions in the Table 5 headings, namely moderate, severe and very severe, refer to severity scores 3, 4 and 5 respectively. None of the asymptomatic plants tested positive for EACMV alone and no weeds tested positive for EACMV or ACMV alone or as a mixed infection (Table 5).
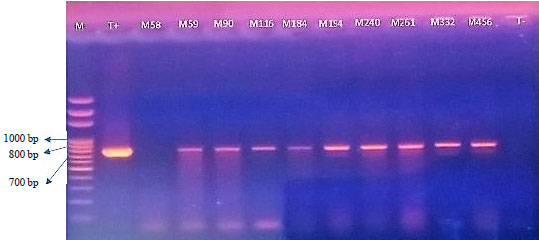 | |
| Fig. 4: | Presence of ACMV |
Lane 1 (M): 1 kb ladder, Lane 2 (T+): Positive control from symptomatic cassava DNA from Burkina Faso, Lane 13 (T-): Negative control (dH2O), Lanes 3-12: Cassava samples M58, M59, M90, M116, M184, M194, M240, M261, M332 and M456, respectively | |
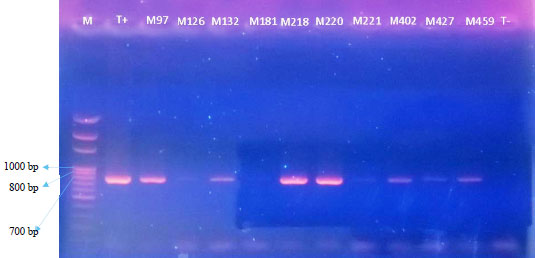 | |
| Fig. 5: | Presence of EACMV |
Lane 1 (M): 1 kb ladder, Lane 2 (T+): Positive control from symptomatic cassava DNA from Burkina Faso, Lane 13 (T-): Negative control (dH2O), Lanes 3-12: Cassava samples M97, M126, M132, M181, M218, M220, M221, M402, M427 and M459, respectively | |
| Table 4: | Incidence (%) by begomovirus in cassava and weed samples |
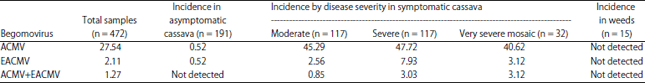 | |
| ACMV: African cassava mosaic virus, EACMV: East African cassava mosaic virus | |
| Table 5: | Incidence (%) by begomovirus in cassava and weeds in 2017 samples |
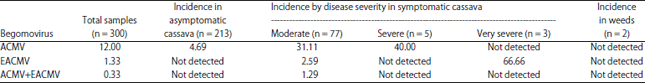 | |
| ACMV: African cassava mosaic virus, EACMV: East African cassava mosaic virus | |
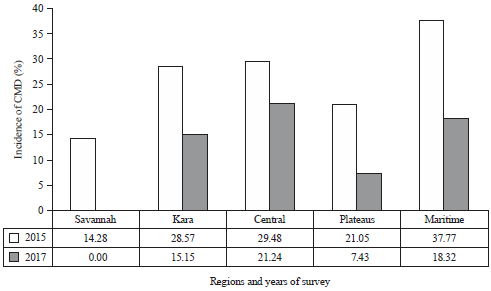 | |
| Fig. 6: | Incidence of CMBs (2015 and 2017) in the 5 agro-ecological regions |
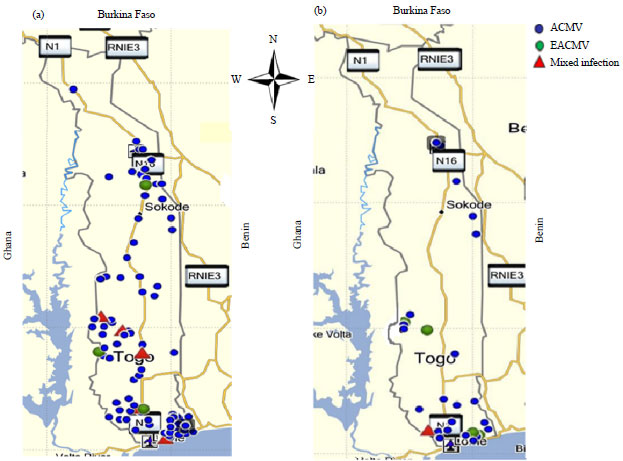 | |
| Fig. 7(a-b): | Distribution of cassava begomoviruses in Togo (a) 2015 and (b) 2017 surveys |
Source: Author’s Work, N1: National Road 1, Lome-Cinkansé, RNIE 3: National highway of Benin, it passes from north to South in the West of the country running near the border with Togo; N16: National Road 16, Kara-Kétao axis leads to the Benin-Togo border to the East | |
Disease incidence by region: In both the 2015 and 2017 surveys, CMD was present in all fields within all regions surveyed (Fig. 6). However, incidence was consistently lower across all regions in 2017 than in 2015.
Single infections of ACMV were recorded in each of the 5 agro-ecological regions. However, single infections of EACMV were less common and were found in the South of the country. Similarly, mixed infections were rare and tended to be distributed in the South of Togo (Fig. 7).
DISCUSSION
The findings of this study confirmed that CMD remains an endemic problem in all agro-ecological zones of Togo. Disease incidence in 2015 varied from one region to another: It was high in Maritime (Southern Togo) and relatively low in Savannah (Northern Togo).
In 2017, disease incidence was high in Central. In contrast, none was recorded in Savannah where only asymptomatic leaves of cassava were collected. The high incidence rates in Maritime and Central during these two survey years suggest that use of infected planting material was probably the cause of the spread of CMD in cassava fields. Farmers have traditionally reused cuttings from their own fields as planting material and these cuttings are often infected with viruses. This leads to the perpetuation of viruses via infected cuttings19. Previous studies reported similar findings for CMD in several countries in sub-Saharan Africa and suggest that the CMD symptoms observed depend on virus species, strains and mixed infections20,21.
Analysis of PCR output for cassava leaf samples, as well as those from symptomatic and asymptomatic weeds, confirmed presence of ACMV and EACMV. The most prevalent begomovirus was ACMV, infecting cassava plants in all cassava-growing areas of Togo22,23. It was found that ACMV incidence rate of 27.54% in 2015 and 13.33 in 2017. Similar observations were reported in Uganda24 and Kenya25. In 2015, the incidence rates of EACMV (2.11 and 1.33%) were higher than those for mixed infections with ACMV, which were 1.27 and 0.33%, in 2015 and 2017, respectively. Previous research showed that ACMV and EACMV were present in infected plants in Africa, either alone or as mixed infections21,26. It has also been shown that inoculum sources of begomoviruses can be traced to naturally infected plants when used as planting material in successive years, as well as to alternative hosts (e.g., weeds)27.
Recombination occurs frequently due to the co-existence of multiple CMB species in one geographical area28. The generation and emergence of highly fit and new recombinant variants can affect cassava productivity: This was the case for the emergence of the severe strain in Uganda called EACMV-UG9. More recently, Kyallo et al.29 proposed that mixed infections may be involved in viral recombination.
In 2015 and 2017, 70.34 and 86.66% of the samples were negative when tested with the primer pairs used in this study, respectively. This suggests that the use of other molecular methods such as rolling circle amplification (RCA) may be necessary to explain this result. Negative results for weeds indicate that these begomoviruses might not be present in these plants. However, Ogbe et al.14 showed that ACMV and EACMV could infect weeds such as Senna occidentalis L. (Fabaceae), Combretum confertum Lams. (Combretaceae) and Ricinus communis L. ( Euphorbiaceae). Therefore, a deeper investigation of all isolates in this study (including weeds) is required: in addition to using RCA, sequencing is required to characterize the isolates and confirm the PCR and RCA assays.
Some asymptomatic cassava leaf samples were positive for ACMV (0.52% in 2015 and 4.69% in 2017) and EACM (in 0.52% in 2015) both survey years. This indicates that absence of visual symptoms on leaves cannot be interpreted as absence of viral infection in the plant.
Based on the surveys of 2015 and 2017, CMD was the main constraint affecting cassava cultivation. Highly effective CMD management programs have been implemented in East Africa to mitigate this disease30,31. These programs are based on the dissemination and multiplication of CMD-resistant cultivars for use as planting material, combined with a rigorous phytosanitary program. At present, no such measures are in place in Togo to address spread of CMD. If the effective results in East Africa are to be replicated in Togo, then concerted efforts will be needed in the immediate future to combine the distribution of healthy cassava planting material with training and extension programs relating to cassava viral diseases.
CONCLUSION
It was found that CMD exists across all cassava-growing regions of Togo, where CMB incidence reached 37.77% (Maritime) in 2015 and 21.24% (Central) in 2017 and the dominant viral strain causing the disease was ACMV. It was considered that implementation of phytosanitation programs for CMD-resistant cassava cultivars are essential to ensure that planting material is CMD-free. This would reduce transmission of CMD in cassava-producing areas of the country. Our study can inform strategies designed to manage this disease in Togo.
SIGNIFICANCE STATEMENT
The research systematically explains the role of CMBs in maintaining the cassava viral epidemic in TOGO. This article will help researchers to find ways to effectively control this viral disease (CMD) in cassava growing fields in Africa and particularly in Togo. Thus a new avenue might be explored scientifically to develop many new resistant plants.
ACKNOWLEDGMENTS
We thank the West African Virus Epidemiology for Togo (WAVE-UL) project funded by the Bill and Melinda Gates Foundation (Grant number OPP1082413) and the UK Department for International Development (DFID) for supporting this project.
REFERENCES
- Maruthi, M.N., J. Colvin, S. Seal and J.M. Thresh, 2002. First report of a distinct begomovirus infecting cassava from Zanzibar. Plant Dis., 86: 187-187.
CrossRefPubMedDirect Link - Fauquet, C.M., R.W. Briddon, J.K. Brown, E. Moriones, J. Stanley, M. Zerbini and X. Zhou, 2008. Geminivirus strain demarcation and nomenclature. Arch. Virol., 153: 783-821.
CrossRefDirect Link - Owor, B., J.P. Legg, G. Okao-Okuja, R. Obonyo and M.W. Ogenga-Latigo, 2004. The effect of cassava mosaic geminiviruses on symptom severity, growth and root yield of a cassava mosaic virus disease-susceptible cultivar in Uganda. Ann. Applied Biol., 145: 331-337.
CrossRefDirect Link - Tiendrebeogo, F., P. Lefeuvre, M. Hoareau, V.S. Traore and N. Barro et al., 2009. Occurrence of East African cassava mosaic virus-Uganda (EACMV-UG) in Burkina Faso. Plant Pathol., 58: 783-783.
CrossRefDirect Link - Akinnagbe, O.M., 2010. Constraints and strategies towards improving cassava production and processing in Enugu North Agricultural Zone of Enugu State, Nigeria. Bangladesh J. Agric. Res., 35: 387-394.
CrossRefDirect Link - Kombate, K., C.P. Kpade, M. Edah, E.L.Y. Loko, R.A.F. Adjatin and A.D. Anagonou, 2016. [Preferences and choice factors of cassava (Manihot esculenta Crantz) varieties in Togo]. Bull. Rech. Agron. Bénin, 2016: 20-29, (In French).
Direct Link - Sseruwagi, P., W.S. Sserubombwe, J.P. Legg, J. Ndunguru and J.M. Thresh, 2004. Methods of surveying the incidence and severity of cassava mosaic disease and whitefly vector populations on cassava in Africa: A review. Virus Res., 100: 129-142.
CrossRefPubMedDirect Link - Ogbe, F.O., A.G.O. Dixon, J.A. Hughes, O.J. Alabi and R. Okechukwu, 2006. Status of cassava begomoviruses and their new natural hosts in Nigeria. Plant Dis., 90: 548-553.
CrossRefDirect Link - Hahn, S.K, E.R. Terry and K. Leuschner, 1980. Breeding cassava for resistance to cassava mosaic disease. Euphytica, 29: 673-683.
CrossRefDirect Link - Permingeat, H.R., M.V. Romagnoli, J.I. Sesma and R.H. Vallejos, 1998. A simple method for isolating DNA of high yield and quality from cotton (shape Gossypium hirsutum L.) leaves. Plant Mol. Biol. Rep., 16: 89-89.
CrossRefDirect Link - Pita, J.S., V.N. Fondong, A. Sangare, G.W. Otim-Nape, S. Ogwal and C.M. Fauquet, 2001. Recombination, pseudorecombination and synergism of geminiviruses are determinant keys to the epidemic of severe cassava mosaic disease in Uganda. J. Gen. Virol., 82: 655-665.
CrossRefDirect Link - Zhou, X., Y. Liu, L. Calvert, C. Munoz, G.W. Otim-Nape, D.J. Robinson and B.D. Harrison, 1997. Evidence that DNA-A of a Geminivirus associated with severe cassava mosaic disease in Uganda has arisen by interspecific recombination. J. Gen. Virol., 78: 2101-2111.
CrossRefDirect Link - Fondong, V.N., J.S. Pita, M.E.C. Rey, A. de Kochko, R.N. Beachy and C.M. Fauquet, 2000. Evidence of synergism between African cassava mosaic virus and a new double-recombinant geminivirus infecting cassava in Cameroon. J. Gen. Virol., 81: 287-297.
CrossRefDirect Link - Ogbe, F.O., G. Thottappilly, A.G. Dixon, G.I. Atiri and H.D. Mignouna, 2003. Variants of East African cassava mosaic virus and its distribution in double infections with African cassava mosaic virus in Nigeria. Plant Dis., 87: 229-232.
CrossRefDirect Link - Adjata, K.D., E. Muller, M. Aziadekey, Y.M.D. Gumedzoe and M. Peterschmitt, 2008. Incidence of cassava viral diseases and first identification of East African cassava mosaic virus and Indian cassava mosaic virus by PCR in Cassava (Manihot esculenta Crantz) fields in Togo. Am. J. Plant Physiol., 3: 73-80.
CrossRefDirect Link - Dansou-Kodjo, K.A., A.S. Mivedor, D.K. Adjata, J. Duclercq, E. Muller and Y.M.D. Gumedzoe, 2017. [Diagnosis of begomoviruses associated with cassava (Manihot esculenta Crantz)-based cropping systems by PCR (Polymerase Chain Reaction) in Togo]. J. Rech. Sci. Univ. Lome, 19: 73-84, (In French).
Direct Link - Harrison, B.D., X. Zhou, G.W. Otim-Nape, Y. Liu and D.J. Robinson, 1997. Role of a novel type of double infection in the geminivirus-induced epidemic of severe cassava mosaic in Uganda. Ann. Applied Biol., 131: 437-448.
CrossRefDirect Link - Bull, S.E., R.W. Briddon, W.S. Sserubombwe, K. Ngugi, P.G. Markham and J. Stanley, 2006. Genetic diversity and phylogeography of cassava mosaic viruses in Kenya. J. Gen. Virol., 87: 3053-3065.
CrossRefDirect Link - Alabi, O.J., F.O. Ogbe, R. Bandyopadhyay, P.L. Kumar, A.G.O. Dixon, J. d’A. Hughes and R.A. Naidu, 2008. Alternate hosts of African cassava mosaic virus and East African cassava mosaic Cameroon virus in Nigeria. Arch. Virol., 153: 1743-1747.
CrossRefDirect Link - Kyallo, M., E.M. Ateka, P. Sseruwagi, J.T. Ascencio-Ibanez, M.O. Ssemakula, R. Skilton and J. Ndunguru, 2017. Infectivity of Deinbollia mosaic virus, a novel weed-infecting begomovirus in East Africa. Arch. Virol., 162: 3439-3445.
CrossRefDirect Link - Legg, J.P., 1999. Emergence, spread and strategies for controlling the pandemic of cassava mosaic virus disease in East and Central Africa. Crop Protect., 18: 627-637.
CrossRefDirect Link